Genesis 1:20-31 God’s Fifth and Sixth Creation Days
“Then God said ‘let the waters team with swarms of living creatures, and let birds fly above the Earth in the open expanse of the Heavens’ (1:20). And God created the great sea monsters and every living creature that moves, with which the waters swarmed after their kind, in every winged bird after its kind; and God saw that it was good (1:21). And God blessed them, saying ‘be fruitful and multiply, and fill the waters in the seas, and let birds multiply on the Earth’ (1:22). And there was evening and there was morning, a fifth day” (1:23).
Il Tintoretto 1518 – 1594 (God’s Creation of the Animals) (1550)1
“Then God said, ‘let the Earth bring forth living creatures after their kind: cattle and creeping things and beasts of the Earth after their kind’; and it was so (1:24). And God made the beasts of the Earth after their kind and the cattle after their kind, and everything that creeps on the ground after its kind; and God saw that it was good (1:25). Then God said, ‘let Us make man in Our image, according to Our likeness; and let them rule over the fish of the sea and the birds of the sky and over the cattle and over all the Earth, and over every creeping thing that creeps on the Earth’ (1:26). And God created man in His own image in the image of God, He created male and female, He created them (1:27). And God blessed them; and God said to them, ‘be fruitful and multiply and fill the Earth and subdue it; and rule over the fish of the sea and over the birds of the sky, and over every living thing that moves on the Earth’ (1:28). Then God said ‘behold, I have given you every plant yielding seed that is on the surface of all the Earth and every tree which has fruit yielding seed; it shall be food for you (1:29); and to every beast of the Earth and to every bird of the sky and to everything that moves on the Earth which has life I have given every green plant for food’ and it was so (1:30). And God saw all that He had made and behold, it was very good. And there was evening and there was morning, the sixth day” (1:31).
Michelangelo Buonarroti 1475 – 1564 (God’s Creation of Adam) (1508-1512)2


Introduction:
During His fifth and sixth days of creation, God created the first animals and humans. Because He created millions of animals during this time period, His eleven-verse summary was not meant to provide an encyclopedia of every animal that He created. Likewise, because the Bible had to be written for people of all ages, He omitted the animals that went extinct before humans, like the dinosaurs and the hominids. References to these creatures would have had no meaning to people living before the 19th Century. Because mankind is the focus of His account, He only lists the animals that were useful to mankind. With a few exceptions, these correspond with the animals later deemed to be “Kosher” under the God’s dietary laws in the Torah. Although not all animal groups are mentioned, there are several lines of proof of God’s special creation during these days.
First, thousands of years ago, Moses revealed that God created the fish and other kinds of sea creatures before creating the animals. Until the 19th Century, believers had only their faith to defend this claim. But beginning in the early 1840s, geologists discovered that nearly every type of fish or ocean-related phyla of organisms appeared before the animals of the Earth in a geological blink of time. This event is now called the “Cambrian explosion”. For more than 150 years, evolutionists have promised that the alleged transitional species leading to the Cambrian explosion would be discovered. But they never have. Moses’ accurate description of this event is confirmation of the Bible’s accuracy. Second, the appearance of amphibians after fish, the next alleged step in evolution, also provides evidence of God’s intelligent design. At a genetic level, there is no evidence that amphibians evolved into fish. Third, the appearance of reptiles after amphibians, the next alleged step in evolution, also provides evidence of God’s intelligent design. The component parts of the first reptilian egg were “irreducible complex” at the time of its first appearance. Moreover, no alleged transitional species between amphibians and reptiles have ever been found. Fourth, the Bible claims that winged animals appeared after the first sea life and before the first land mammals. Again, scientists have confirmed the Bible on this point to be accurate. Fifth, the appearance of the first warm-blooded mammals, the next alleged step in evolution, also provides evidence of God’s intelligent design. The internal systems of the first warm-blooded mammal were also irreducibly complex at its first appearance. Moreover, with only a few disputed exceptions, evolutionists do not have any transitional fossils for the nearly 4,000 types of mammals alive today. Sixth, the next alleged evolutionary step in the appearance of mankind is the primate. But after more than 150 years of searching, scientists have never found anything that primates allegedly evolved from. There is likewise no evidence that withstands the test of scrutiny that they or the hominids evolved into humans. Finally, thousands of years ago, Moses claimed that mankind was God’s final creation. For centuries, believers had nothing but their faith to defend this claim. With the exception of new strands of viruses or bacteria, scientists have verified that no new species of animals have appeared since mankind. This again shows the Bible to be God’s inspired Word. The Bible also proclaims that God made mankind in His image. The unique attributes of mankind distinguish it from anything that preceded it. Among other things, humans alone have the ability to concern themselves with questions of good and evil and morality. The unique aspects of humanity contain the fingerprints of God’s creation. These multiple lines of evidence confirm both God’s special creation during His fifth and sixth days and the Bible’s authenticity.
The following chart is a chronology of the first appearance of every major organism or animal group in the alleged evolutionary sequence from bacteria to mankind. The prior study for God’s third and fourth creation days established that the first bacteria and the first “protist” (a eukaryotic celled organism) miraculously appeared with no predecessor organisms. Moreover, the hostile conditions of the early Earth would not allow for organic materials to form on their own. Modern theories of evolution further offer no support for the natural origin of life. God’s fifth and sixth creation days also bear His finger prints through multiple progressive acts of special creation until the final appearance of mankind.
God’s Progressive Creation of Bacteria to Mankind (Creation Days Three Through Six)
| GOD’S CREATION DAY/ TYPE OF ORGANISM | ERAS | PERIODS | EST. TIME SPAN, YEARS AGO |
|---|---|---|---|
(Third and Fourth Days) Earth cools Oceans appear Fully formed anaerobic bacteria appear At least 30 life-exterminating asteroid impacts on Earth “cyanobacteria”; Blue green algae - prokaryotic bacteria; Protists (eukaryotic celled); Photosynthetic land plants |
Archeozoic Proterozoic |
4.54 billion years ago 4.41 billion years ago 4.28 billion years ago.3 From 3.8 until 3.5 billion years ago.4 Between 2.7 billion to 2.1 billion ago.5 3 billion to 1.7 billion years old.6 730,000,0007 |
|
(Fifth and Sixth Days) (“Cambrian Explosion”; Invertebrates; jawless fish; jawed fish; boney lobe-finned fish; (Gen. 1:20-21.) Amphibians; |
Paleozoic Mesozoic |
Cambrian Triassic |
540,000,0008 370 million years ago.9 |
Reptiles; Avian dinosaurs / birds (Gen 1:22-23) Mammals (Gen. 1:24-25.); Primates |
Mesozoic | Carboniferous Jurassic Cretaceous |
320 to 310 million years ago10 240 million years ago 200 to 125 million years ago.11 65 million years ago.12 |
Humans (Gen. 1:26-28; 2:7.), The cultural “Big Bang.” |
Recent | Pleistocene Epoch | 200,000 to 50,000.13 50,000 years ago.14 |
1. The “Cambrian Explosion”: The Bible’s Accurate Description of God’s Creation of the Fish and Other Sea Life During His Fifth Creation Day. Gen. 1:20-23.
Thousands of years ago, Moses made a bold claim. He alleged that God made the fish after the first plant life and before the animals or humans. In the Genesis account, he specifically names fish, which are Kosher under God’s dietary laws. By referencing “swarms” or “sheres” (Gen. 1:20), he also makes a unique reference to the non-Kosher organisms of the sea, like mollusks and crustaceans. Furthermore, through the use of two different Hebrew words for “create”, Moses further reveals that God created some sea life from nothing (“bara”) and others using existing resources (“asa”).15 Thus, for some organisms, God created them from nothing. For others, He most likely created new organisms by guiding change within the embryo of the organism.
Evolutionists often claim that the creationists can only raise negative argumentation to prove their points. But Moses’ amazing description of all the fish and ocean-related phyla appearing at once and before the land animals has now been confirmed.
The Bible’s accuracy and God’s intelligent design is evidenced by at least seven things. This include: (1) the sudden appearance of nearly every known phyla of organisms (something which is consistent with the Bible and contrary to any accepted theory of evolution); (2) the absence of any intermediate organisms, (despite more than 150 years of searching); (3) the appearance of different kinds of eyes and complex organs (all without any predecessor organisms); (4) the inability for large predecessor organisms to have existed based upon the low pre-Cambrian oxygen levels in the atmosphere and a global ice age that preceded the Cambrian explosion; (5) the inability of more modern theories of evolution, like punctuated equilibrium, to explain this explosion of organisms; (6) the inability of the theories of genetic mutation to explain this explosion of organisms; and (7) the impossibility of every possible predecessor organisms staying hidden in the fossil record after more than 150 years of searching.
(1) The Sudden Appearance of Every Known Animal Phyla.
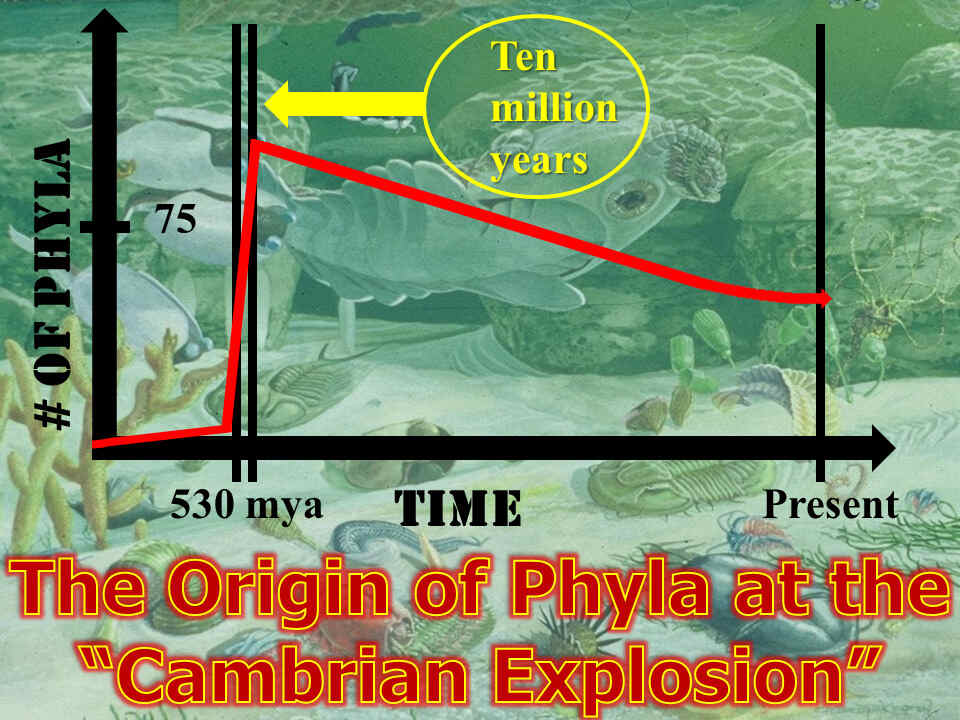
Approximately 540 million years ago during the Paleozoic era, every animal phyla ever to exist, between 70 and 75 in total, suddenly appeared. (A phylum is as groupings of animals based upon their general body plan. A phylum is the highest level in a system for classifying organisms, which a Swedish botanist, physician, and zoologist named Carl Linnaeus set up in 173516). This further occurred in a geological “blink of an eye,” spanning between 5 to 10 million years. Since that time, no new animal phyla have arisen. By some estimates, the number has in fact decreased.
Within this short time, fossils abound of fully formed complex invertebrates. This also includes “chordates”, to which vertebrates (animals with backbones) such as humans belong.
The animals included “brachiopods.” These creatures lived in shells that looked like those used by clams or cockles.
The animals further included “arthropods.” These are animals with jointed, external skeletons. Evolutionists call them the ancestors of spiders and crustaceans and certain insects. Their bodies also showed that complex predator-prey relationships existed when no predator-prey relationships previously existed. Their hard exteriors offered both a defense against enemies and a framework for supporting larger body sizes.
The animals also included “trilobites.” To protect themselves in the oceans from predators that also instantly appeared during this time period, they had flattened, segmented, plated bodies. They also appeared with widely varying sizes. Some were only a millimeter in length. Others were more than 2 feet in length.
Other animals included the “Anomalocaris.” This animal looks like a giant shrimp. Its mouth had hooks to kill its prey.
These animals all had complex eyes or organs when they first appeared. For example, the “Opabinia” had five eyes. Along with a flexible clawed arm attached to its head, it used these five eyes to catch its prey.
The diverse animals include, but not limited to: clams, worms, archaic sponges, sea urchins, sea lilies, snails, jellyfish, brachiopods, sea cucumbers, swimming crustaceans, and other complex invertebrates. Every single one appeared fully formed and without any trace of prior transitional species.
The amazement of this explosion of life is not limited to the sudden appearance of all 70 to 75 animal phyla and a variety of fully formed invertebrates. Many of the major insect groups known today also appeared in the fossil record, some as we see them today.
The Cambrian fossil record also includes a high number of “lagerstätten.” These are specially occurring geological events that preserve soft tissue. These lagerstätten have allowed paleontologists to examine the internal anatomy of animals, where, in other sediments, only shells, spines, and claws remain. The most significant Cambrian lagerstätten are found in the Maotianshan shale beds of Chengjiang in Yunnan, China; the Sirius Passet in Greenland; the Burgess Shale in British Columbia, Canada; and in the Orsten fossil beds of Sweden. From these lagerstätten, paleontologists have verified that many of these organisms, like the snails, crustaceans, and insects, had fully formed eyes and digestive organs.17
Discoveries of marine crustacean fossils from the Maotiansham Shale of China further reveal evidence of the simultaneous appearance of open water (pelagic) animals, fully equipped with specialized swimming organs, eyes, and defensive shells, at the same time as bottom dwelling (benthic) animals appeared. Under an evolutionary model, open water organisms should have evolved long after the appearance of bottom dwelling organisms.18
This amazing explosion of invertebrates throughout the entire world has been popularly termed by geologists the “Cambrian explosion.” Others call it life’s “Big Bang.”
The amazement of the Cambrian explosion does not end with these discoveries. Other discoveries show that two entire groups, previously assumed to represent the next steps in our evolutionary pathway, also appeared at the same time and without a trace of intermediates.
The first discovery includes fossilized fish in the middle of the Cambrian rocks.19 Examples include the soft spinal cord cordobite fish called the “Pikaia gracilens.” It was a wormlike creature that swam in the Cambrian seas. Fossils found in the Burgess Shale of British Columbia show traces of a notochord (a rod-like primitive backbone).20
Until the late 1990s, paleontologists upheld this organism with its complex dorsal notochord as the transitional species linking invertebrates to hard spinal cord vertebrates. Paleontologists, however, have radiocarbon dates of this organism showing its existence approximately 535 million years ago. This places the Pikaia right in the beginning to middle of the Cambrian explosion when other invertebrates first appeared.21 Moreover, this organism had fully developed eyes, and it is believed that it burrowed its tail into the sand and fed off of microscopic food.
In 1999, Chinese paleontologists also found two different kinds of jawless, boney lobbed fish in the 530-million-year-old Chengjiang fossil bed in China. One is called “haikouichthy.” It looks just like a modern fish. The boney lobed fish were presumed to be the next step on the evolutionary pathway to humans. Like the other organisms discovered during this period, these fish had fully formed complex organs like eyes and digestive organs.22
Hugh Ross observes that the Cambrian explosion further contained an astonishing 146 of 182 possible skeletal designs: “The fossil record testifies that for the first 85 percent of life’s history on Earth there were no creatures we would refer to as animals. Then, suddenly, in shallow seas and on continental shelves, life-forms manifesting nearly every conceivable body plan appeared. In both the Avalon explosion (575 million years ago) and the Cambrian explosion (543 million years ago) swarms of diverse sea animals abruptly emerged. For the first time in Earth’s history creatures sported appendages, limbs, skeletons, and specialized organs. In fact, of the 182 mathematically possible skeletal designs conceivable for physical life, 146 show up in the fossil records of the Cambrian explosion event.”23
Thus, the Bible made an accurate claim thousands of years ago that can be tested against the fossil record. If the Bible were simply an invented book of myths, Moses could not have made these types of accurate predictions about the fossil record. Thus, the fossil record confirms that the Bible is the inspired Word of God with information that no human could have known or independently guessed. “All Scripture is inspired by God and profitable for teaching, for reproof, for correction, for training in righteousness;” (2 Tim. 3:16).
(2) The Absence of Any Intermediate Organisms.
Most advocates of evolution vehemently dispute that God had anything to do with this explosion of life. But the 70 to 75 animal phyla that appear during the Cambrian explosion also appear without a single transitional organism preceding them.
The only multicellular life that preceded the Cambrian explosion (that was not a plant) was the “Ediacara.” (see picture below) This organism appeared in what is called the “Avalon explosion”, 575 million years ago.24 “Ediacarans were similar to plants. They were immobile, tube-shaped creatures that didn’t eat each other.”25 These organisms were so different from anything that preceded them or followed them that some paleontologists believe that they were in a separate kingdom level category “Vendozoa” (now renamed “Vendobionta”).26 Thus, these organisms were not transitional to anything that appeared during the Cambrian explosion.

In 1841, scientists first documented this explosion of fossils in the Cambrian era rocks.27 Although exploration of the Cambrian rocks and scientific knowledge of these creatures preceded 1841, scientists at that time began to earnestly investigate this phenomena.
In 1859, 18 years later, Darwin conceded that scientists could find no evidence of any transitional species before this explosion: “There is another and allied difficulty, which is much more serious. I allude to the manner in which species belonging to several of the main divisions of the animal kingdom suddenly appear in the lowest known fossiliferous rocks.28 . . . The abrupt manner in which whole groups of species suddenly appear in certain formations, has been urged by several paleontologists—for instance, by Agassiz, Pictet, and Sedgwick—as a fatal objection to the belief in the transmutation of species. If numerous species, belonging to the same genera or families, have really started into life at once, the fact would be fatal to the theory of evolution through natural selection.29 …”
In the sixth edition of his book, he again conceded: “To the question why we do not find rich fossiliferous deposits belonging to these assumed earliest periods prior to the Cambrian system, I can give no satisfactory answer.30 . . . The case at present must remain inexplicable, and may be truly urged as a valid argument against the views here entertained.”31
For more than 150 years, paleontologists have scoured the fossil record in the most remote places of the Earth searching for the transitional species that had to exist for Darwin’s theory to be true. But paleontologists have never found a single transitional species dated prior to life’s “Big Bang”.
Faced with this reality, well-known modern paleontologists have conceded that these transitional organisms (which are necessary for Darwin’s theory to be true) simply cannot be found. Many of the following quotes come from the late biochemist Dr. Duane Gish (1921-2013).
For example, the late Daniel I. Axelrod (1910 to 1988) conceded before his death: “One of the major unsolved problems of geology and evolution is the occurrence of diversified, multicellular marine invertebrates in Lower Cambrian rocks on all the continents and their absence in rocks of greater age.”32
Professor of Geology T. Neville likewise concedes: “Granted an evolutionary origin of the main groups of animals, and not an act of special creation, the absence of any record whatsoever of a single member of any of the phyla in the Precambrian rocks remains as inexplicable on orthodox grounds as it was to Darwin.”33
Geologist and paleontologist James W. Valentine likewise concedes: “The fossil record is of little use in providing direct evidence of the pathways of descent for phyla or invertebrate classes. Each phyla within the fossil record had already evolved its characteristic body plan when it first appeared. So far as we can tell from the fossil remains, no phylum is connected to any other via intermedia fossil types. Indeed none of the invertebrate classes can be connected with another class by a series of intermediates . . . Each of the phyla that developed durably skeletonized lineages during this period did so independently, suggesting that the opportunities for epifaunal life were open to a wide array of adaptive types. Furthermore, many of the durably skeletonized phyla appearing in the Cambrian rocks are represented by a number of distinctive subgroups, classes, or orders that appear suddenly without known intermediates.”34
Swedish paleontologist Stephen Bengtson likewise concedes: “If any event in life's history resembles man’s creation myths, it is this sudden diversification of marine life when multicellular organisms took over as the dominant actors in ecology and evolution. Baffling (and embarrassing) to Darwin, this event still dazzles us and stands as a major biological revolution on a par with the invention of self-replication and the origin of eukaryotic cell. The animal phyla emerged from the pre-Cambrian mist with most of their attributes of their modern descendants.”35
Others concede that the fossil record provides no support in explaining this explosion of life: “Most of the animal phyla that are represented in the fossil record first appear, ‘fully formed’ and identifiable as to their phylum in the Cambrian some 550 million years ago...The fossil record is therefore of no help with respect to the origin and early diversification of the various animal phyla…”36
As stated above, the allegedly transitional fish have now also been found together in the Cambrian explosion. Even before the discovery of these fish in the Cambrian sedimentary rocks, a renowned paleontologist, the late Alfred Romer (1894 – 1973), conceded before his death that there are simply no transitional species in the alleged fossil record leading to the first appearance of fish: “In sediments of late Silurian and Devonian age [now found in the Cambrian explosion], numerous fish-like vertebrates of varied type are present, and it is obvious that a long evolutionary history had taken place before that time. But of that history we are mainly ignorant.”37
Regarding the first appearance of bony-fish, Romer conceded that their appearance in the fossil record is a “dramatically sudden one.”38 Separately, he conceded: “The common ancestor of the bony-fish groups is unknown. There are various features, many of them noted above, in which two typical subclasses of bony fish are already widely divergent when we first see them . . .”39
Another prominent evolutionist likewise questions how they could have originated without a trace of any transitional or prior similar organism: “All three subdivisions of the bony fishes appear in the fossil record at approximately the same time. They are already widely divergent morphologically, and they are heavily armored. How did they originate? What allowed them to diverge so widely? How did they all come to have heavy armor? And why is there no trace of earlier intermediate forms?”40
Evolutionist Strahler likewise concedes that the: “origin of the vertebrates is obscure - there is no fossil record preceding the occurrence of fishes . . ..”41 He then conceded: “This is one count in the Creationists’ charge that can only evoke in unison from the paleontologists a plea of nolo contendere.”42
The plea of nolo contendere is a plea of a defendant in a court proceeding who can offer no defense. It is like waving the white flag of surrender.
Even more astonishing, a prominent evolutionist and expert on fishes, Errol White, once openly conceded during a speech regarding lungfishes: “But whatever ideas authorities may have on the subject, the lungfishes, like every other major group of fishes that I know, have their origins firmly based in nothing . . . I have often thought of how little I would like to have to prove organic evolution in a courtroom of law.”43
Professor Jeffrey S. Levinton also asks why all of life’s body plans appeared at once with no new ones continuing to be developed. Indeed, by some estimates, the total number of phyla have actually decreased since the Cambrian explosion: “Evolutionary biology’s deepest paradox concerns this strange discontinuity. Why haven’t new animal body plans continued to crawl out of the evolutionary cauldron during the past hundreds of millions of years? Why are the ancient body plans so stable?”44
(chart source45)
Richard Dawkins, an Oxford evolutionary biologist, who frequently attacks both religion and those who believe in God’s special creation, also concedes that there is absolutely no evidence of any transitional species before the Cambrian explosion: “. . . the Cambrian strata of rocks, vintage about 600 million years ago [evolutionists are now dating the beginning of the Cambrian at about 540 million years] are the oldest in which we find some of the major invertebrate groups. And we find many of them already in an advanced state of evolution, the very first time they appear. It is as though they were just planted there without any evolutionary history. Needless to say, this appearance of sudden planting has delighted creationists.”46
Of course, if Dawkins were impartial and objective, he would follow the science wherever it leads. He would not express disappointment when it points towards God.
(3) The Appearance of Multiple Kinds of Fully Formed Eyes and Organs.
In addition to the appearance of between 70 and 75 animal phyla (i.e., body plans) without any transitional species; insects, invertebrates, cordobites, and boney lobed fish all had different kinds of fully functioning eyes when they first appeared. These multiple different kinds of eyes further cannot be explained through prior organisms with more primitive eyes. With the one unrelated exception of the Ediacara, only single celled organisms existed before the Cambrian explosion. Further, these different types of eyes could not have evolved from a single common ancestor just before the Cambrian explosion. An insect eye is radically different from the eyes of invertebrates and vertebrates. Thus, for Darwin’s theory to be true, there needed to be multiple kinds of different transitional organisms with different kinds of predecessor eyes. But they simply do not exist.
For example, in the Emu Bay Shale of South Australia, researchers found a 515 million-years-old compound eye, like a modern fly (see below.) It was the corneas of an arthropod called “Anomalocaris” (see below). From this eye, researchers have found that: “the earliest arthropods possessed highly advanced compound eyes.” For one recovered specimen, researchers found that each eye had “over 3,000 large ommatidial lenses.” This allowed for extremely advanced sight: “The arrangement and size-gradient of lenses creates a distinct [forward] ‘bright zone’... where the visual field is sampled with higher light sensitivity (due to larger ommatidia) and possibly higher acuity.” This indicates “that these eyes belonged to an active predator that was capable of seeing in low light.” These eyes were further very different from the trilobites at that time: “The eyes are more complex than those known from contemporaneous trilobites and are as advanced as those of many living forms” today.47
A compound eye of a modern fly is magnified below to show the specialized ommatidial lenses used by arthropods in the past:48

The compound eye was found in the Cambrian arthropod below called Anomalocaris, a three-foot long sea predator:49
 |
|---|
By contrast, the “opabinia” below further had five different eyes on it, like some modern spiders:50
 |
|---|
By contrast, the “haikouichthy” below also appeared 530 million years ago with fish eyes:51

Darwin admitted that if any complex organ could be shown to have always existed his theory would be disproved. In The Origin of Species (1859), he conceded: “To suppose that the eye [...] could have been formed by natural selection, seems, I freely confess, absurd in the highest degree.” But he maintained faith that his theory would one day provide an answer.
Likewise, Dawkins concedes: “The idea of a single macromutation giving rise to a fully functioning eye with the properties listed above, where there was only bare skin before, is, indeed, just about as improbable as a hurricane assembling a Boeing 747.”52
The common response amongst evolutionists is to claim that everything is possible with large periods of time. But with the one exception of the multi-celled Ediacara (which is in a different animal kingdom) only single celled organisms preceded the Cambrian explosion.
Thus, life did not have a long period of time for these different kinds of eyes and specialized organs to appear. Again, these specialized organs and completely different kinds of eyes all appeared in a geological blink of the eye. By the absence of any processor organs leading up to these different kinds of eyes, Darwin leads us to an inescapable conclusion as to the soundness of his theory. The full appearance of these complex organisms and organs disproves his theory. Each complex eye and organ bears the fingerprint’s of God’s intelligent design. “The Creator's wisdom and power are to be admired as much in an ant as in an elephant.” (Matthew Henry on Gen. 1:20-25).53
(4) The Inability of Predecessor Organisms to Have Existed Based Upon the Earth’s Precambrian Oxygen Levels and the Prior Global Ice Age.
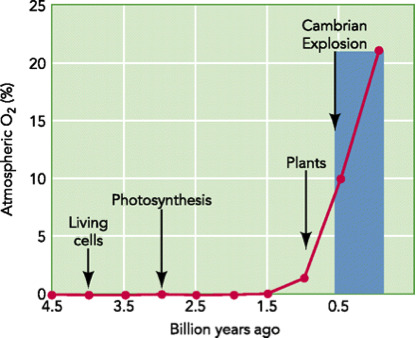
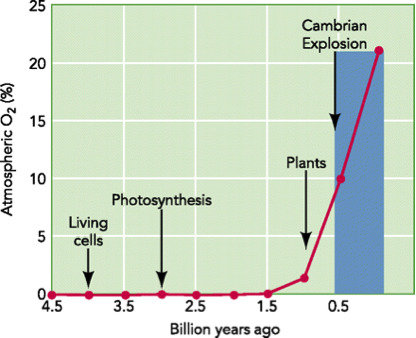
For several reasons, the Precambrian environment could not have supported large predecessor organisms. Soon after the Earth formed, oxygen existed in the early Earth atmosphere in only a limited amount from volcanic gases. The amount of oxygen was enough to break down organic matter, like proteins or amino acids, that was not protected within the first living bacteria. But there was not enough oxygen to sustain multi-celled life. Most of the bacteria thriving on Earth were “anaerobic”. This means that they metabolized their food without oxygen.
At that time, most of Earth’s oxygen was bound up with water molecules or bound to iron on the Earth. But between 2.7 billion to 2.1 billion years ago, God introduced the first “cyanobacteria”, blue-green algae in the oceans. These are prokaryotic bacteria that still exist today.54 For millions of years, blue-green algae produced oxygen as a waste product while converting sunlight into energy. Later types of photosynthetic plant life also produced oxygen.55
This oxygen was, however, toxic to the first anaerobic bacteria. As the oxygen levels slowly increased, this led to the Earth’s first documented mass extinction event. Most (but not all) of the anaerobic bacteria were wiped out by the production of this oxygen. This mass extinction of anaerobic bacteria in turn allowed the amount of oxygen in the Earth’s atmosphere to rapidly increase. Anaerobic bacteria became confined to the low oxygen places of the Earth, like the bottom of the ocean.
This explosion of oxygen, however, did not begin until just before the Cambrian explosion. As soon as the oxygen levels were high enough to sustain life’s first sea creatures, they appeared in the geological blink of the eye. Geologists call this “the Great Oxidation Event.”
The absence of significant oxygen before the Cambrian explosion explains why geologists have not been able to find transitional organisms before the Cambrian explosion. Simply put, without significant oxygen, these organisms could not have existed.
(chart source56)
Hugh Ross also points out three additional reasons why complex organisms could not have appeared sooner. First, the shallow seas and continental shelves needed for the Cambrian life to exist did not appear from tectonic activity and erosion until this point in the Earth’s history. Second, the Sun’s luminosity was slowly increasing to the point where advanced life could exist. Third, just before the Cambrian explosion, most of the Earth was nearly frozen over by a climatic event that would have wiped out most life on the planet: “Runaway freeze events have covered up to 90 percent of Earth’s surface area in an icy blanket. One such snowball event is known to have occurred just prior to the Avalon and Cambrian explosions.”57
For more than 150 years, scientists have argued as a matter of faith that the missing transitional forms will be found at a later time. But the full fossil record, the oxygen record, the Earth’s geological record, the Sun’s luminosity record and the ice age before the Cambrian explosion together establish that these transitional organisms are missing because they could not have existed.
Moses had no access to the fossil record. He did not know the Earth’s oxygen record. He further would have no way of knowing if fish appeared before animals and humans or at the same time. His writings provide a testable model to show that the Bible was written by a source beyond human knowledge at the time.
(5) The Inability of Modern Theories of Evolution, Like Punctuated Equilibrium, to Explain the Cambrian Explosion.
(a) The absence of transitional species in every area of the fossil record.
The absence of transitional organisms is not limited to the Cambrian explosion. This absence of transitional organisms exists throughout the entire fossil record. Speaking broadly regarding the entire fossil record, Darwin posed the question to himself: “Why, if species have descended from other species by insensibly fine predations, do we not everywhere see innumerable transitional forms? Why is not all nature in confusion instead of the species being, as we see them, well defined?”58
Darwin proposed that the gaps between the species could be accounted for by intermediate species long ago extinct. He, nevertheless, conceded that the state of the fossil evidence in his time was lacking in all areas, not just the Cambrian explosion: “‘the most obvious and greatest objection which can be urged against my theory’ and that it accounted for the fact that all the most eminent paleontologists . . . and all our greatest geologists . . . have unanimously, often vehemently, maintained the immutability of species.’”59
Darwin speculated that the number of missing intermediate transitional links between the species, hidden within the Earth, “must have been inconceivably great.”60
Likewise, Thomas Huxley (1825-1895), who was called “Darwin’s bulldog”, placed so much importance upon the theorized transitional species hidden within the Earth that he conceded: “‘If it could be shown that this fact [gaps between widely distinct groups] had always existed, the fact would be fatal to the doctrine of evolution.’”61
By Huxley’s own standard, if he were alive today, he would be forced to concede defeat. Noted paleontologist, the late Stephen J. Gould of Harvard conceded before his death: “‘The fossil record with its abrupt transitions offers no support for gradual change . . . all paleontologists know that the fossil record contains precious little in the way of intermediate forms . . . Transitions between major groups are characteristically abrupt.’”62
In a different publication, he again conceded that: “The absence of fossil evidence for intermediary stages between major transitions in organic design, indeed our inability, even in our imagination, to construct functional intermediates in many cases, has been a persistent and nagging problem for gradualistic accounts of evolution.”63
In a third publication, he again conceded: “The extreme rarity of transitional forms in the fossil record persists as the trade secret of paleontology. The evolutionary trees that adorn our textbooks have data only at the tips and nodes of their branches; the rest is inference, however reasonable, not the evidence of fossils.”64
Before his death, Gould protested bitterly each time a creationist quoted his concessions. But he is not alone in his concessions. The late Dr. George Gaylord Simpson (1902-1984), a Harvard professor in vertebrate paleontology and a noted proponent of evolution, also once conceded: “‘The regular absence of transitional forms is an almost universal phenomenon . . . it remains true, as every paleontologist knows, that most new species, genera, and families and that nearly all new categories above the level of families, appear in the record suddenly and are not led up to by known, gradual, completely continuous transitional sequences’”65
Harvard paleontologist Niles Eldredge, Gould’s former colleague, has also conceded: “No wonder paleontologists shied away from evolution for so long. It never seemed to happen. Assiduous collecting up cliff faces yields … a rate too slow to account for all the prodigious change that has occurred in evolutionary history. When we do see the introduction of evolutionary novelty, it usually shows up with a bang, and often with no firm evidence that the fossils did not evolve elsewhere!”66
Likewise, another noted vertebrate paleontologist Robert Carroll once conceded: “Fossils would be expected to show a continuous progression of slightly different forms linking all species and all major groups with one another in a nearly unbroken spectrum. In fact, most well-preserved fossils are as readily classified in a relatively small number of major groups…”67
Zoologist Mark Pagel also once conceded: “Paleontologists flocked to these scientific visions of a world in a constant state of flux and admixture. But instead of finding the slow, smooth and progressive changes Lyell and Darwin had expected, they saw in the fossil records rapid bursts of change, new species appearing seemingly out of nowhere and then remaining unchanged for millions of years–patterns hauntingly reminiscent of creation.”68
Evolutionary biologist Ernst Mayr (1904 – 2005) likewise conceded before his death: “Wherever we look at the living biota … discontinuities are overwhelmingly frequent…The discontinuities are even more striking in the fossil record. New species usually appear in the fossil record suddenly, not connected with their ancestors by a series of intermediates.”69
And the list goes on and on. Without a fossil record to support a theory of gradual evolution leading up to the Cambrian explosion or the first appearance of any other organisms in the fossil record, evolutionists have sought out other theories to support these unexplainable explosions of life. But these alternative theories also fail.
(b) Punctuated Equilibrium
In 1972, paleontologists Niles Eldredge and the late Stephen Jay Gould published a paper to explain their theory for the gaps in the fossil record. They called it “punctuated equilibria”.70 Speaking broadly and not limited to the Cambrian explosion, they conceded that Darwin’s theory of gradual change in the fossil record at the species level is virtually nonexistent. They instead conceded that “stasis” dominates the history of most fossil species. They alleged that, when significant evolutionary change occurs, it is generally restricted to rare and geologically rapid events of branching speciation called “cladogenesis”. Cladogenesis is the alleged process by which a species splits into two distinct species instead of one species gradually transforming into another. Under punctuated equilibrium, species typically change little as, “gradual change is not the normal state of a species.”71 Also under this theory, large populations may experience, “minor adaptive modifications of fluctuating effect through time.” But they will “rarely transform in toto to something fundamentally new.”72 By contrast, Darwin believed that evolution occurred uniformly and by the steady and gradual transformation of whole lineages, called “anagenesis” or “phyletic gradualism”.
(c) Allopatric Speciation.
To understand where the theory of punctuated equilibrium falls short, a brief explanation is required regarding the difference between observed and agreed upon change in organisms (“micro-evolution”) and the theorized development of new species (“macro-evolution”). The theory of punctuated equilibrium borrows elements from a theory accepted by both evolutionists and creationists called “allopatric speciation.” This is the process of change where members of an individual species or group become isolated from the parent species. Organisms that become separated from the parent species are often forced to interbreed in smaller groups. This process often results in a “genetic drift” in the frequency in which certain genetic traits will appear. When changes in the frequency of genes are extensive enough, one allele of a gene pair can disappear completely from a population. An “allele” is one of the pair of genes that occupy an organism’s chromosomes and determine alternative expressions of a single trait. This can explain why humans developed different color eyes and skin colors after they left from a common place of origin long ago. These changes all result from the loss of genetic information in the sub-group.
(d) The Founder and Bottleneck Effects.
Evolutionists and creationists also agree that reproductive isolation resulting in genetic drift can occur from one of two natural circumstances. These include the “founder effect” and the “bottleneck effect.” The founder effect occurs when organisms of an individual species colonize a geographically distinct area like an island and become separated from the parent species. The bottleneck effect occurs when an environmental event, such as a glacier, causes the separation of members of the same species and a general reduction in the number of organisms available for the breeding pool. The Hawaiian Honeycreeper and the Galapagos Finch provide examples of birds which colonized distinct islands and developed minor changes through separation from the parent species. These birds experienced change due to environmental circumstances. But their bodies and features always show boundaries to how much they can change.
Geographic separation can also result in an offspring that can no longer interbreed if there is a break in a “breeding chain.” In a breeding chain, members within the chain are capable of breeding with members of the same species closest in proximity or size, but not those furthest away. As an example, many small dogs can naturally breed with the mid-sized dogs, but not directly with the largest sized dogs. If all mid-sized dogs were cut off from the largest and smallest dogs, the breeding chain would be broken. The largest and smallest dog would still be dogs. But they would not be able to breed directly with each other.
Likewise, the Helizonius erato butterfly, located in the Amazon and Central American rainforests, provides another example of the “breeding chain” effect. These butterflies live in an immense jungle region, which stretches over an area the size of the European Continent. Over time, numerous sub-populations have developed through the geographic separation of the members of the group at the furthest ends. Although adjacent sub-populations can interbreed with each other, sub-populations found at the opposite edges of the rainforest have less interfertility and could, theoretically, lose their ability to interbreed over time.
(e) The Ambiguous Definition of a Species.
Thus, both evolutionists and creationists agree that allopatric speciation and other related theories document minor changes in organisms. In some cases, these minor changes may result in the inability of parts of a species to interbreed after being separated from the parent group over time. But it is debatable whether any of these minor variations result in a new species. The reason for this uncertainty is that the term “species” is not subject to a strict definition. In relevant part, the New World Dictionary of the American Language (Simon and Schuster 2 ed, 1980) p. 1367 defines a species as a group capable of interbreeding: “The fundamental biological classification, comprising a subdivision of genus and consisting of a number of plants or animals, all of which have a high degree of similarity, can generally interbreed only among themselves, and show persistent differences from members of allied species.”
But the general notion that a species cannot interbreed with other similar members is riddled with contradictions. Commentator Richard Milton provides examples of how this definition does not work in practice. First, the test is not applicable to plants and animals that do not reproduce sexually, such as sea squirts or self-pollinating plants. This is a substantial part of the biological world. Second, the test cannot generally be applied to plants and animals that are extinct and that are known only from their fossils. Again, this is the vast majority of organisms.73
Third, the test leads to some inexplicable anomalies. For example, there are some breeding populations of the (such as the fruit fly Drosophila) that are described as separate species and that do not, (or cannot) interbreed. But they are genetically identical. 74
Fourth, even amongst mammals, the test cannot be applied consistently. For example, any offspring of a horse and an ass (a mule or hinny) should be infertile. But cases have been reported of hinnies bearing offspring. Similarly, the domestic bull Bos taurus can be crossed with a North American Buffalo bison to produce a hybrid, the cattalo, which is fertile. 75 As another example, the Atlantic bottlenose dolphin and the larger pacific dolphin have always been considered different species because they are different in size, live in different places and do not interbreed. But in the 1990s, the two different species of dolphin bred together when placed in a joint training tank of SeaWorld in San Diego California.
These contradictions are important because many evolutions label sub-groups as new species based upon their difficult interbreeding. They then point to this as evidence of macroevolution over long periods of time.
(f) The Limits of Change Observed Through Artificial Selection.
Artificial selection confirms that micro-variations have limits of change. In the case of selective breeding of domestic dogs, humans have successfully bred smaller and larger sized dogs. Nevertheless, breeders of dogs have discovered that, at a certain point in time, dogs reach a genetic limit in their size variation. In other words, artificial breeding of dogs has revealed the impossibility of breeding dogs that continue to grow in size to elephants or as small in size as rats.
With this point, Dawkins agrees, “In a few hundred years, or at the most a few thousand we have gone from wolf to Pekinese, Bulldog, Chihuahua and Saint Bernard. Ah, but they are still dogs aren’t they? Yes, if it's comforting to play with words like that, you can still call them dogs.”76 Dawkins nevertheless argues against the observed limits of change that humans have documented to argue that, with infinite time, we might hypothetically see changes resulting in something that is not a dog. But this theory of what might happen is not proof that macroevolution does happen.
(g) Punctuated Equilibrium’s Claim that It Cannot be Tested.
Punctuated equilibrium imagines that subspecies go into a genetic mutation hyper-drive once they are separated from the main group. As Dawkins explains: “The proponents of punctuated equilibrium . . . believe that there are genetic forces in large populations that actively resist evolutionary change, for them, is a rare event, coinciding with speciation. It coincides with speciation in the sense that, in their view, the conditions under which new species are formed - geographical separation of small, isolated subpopulations - are the very conditions under which the forces that normally resist evolutionary change are relaxed or overthrown. Speciation is a time of upheaval, or revolution. And it is during these times of upheaval that evolutionary change is concentrated. For most of the history of a lineage it stagnates.”77
Dawkins concedes that, when a population is not isolated, interbreeding within the general population prevents change: “To a punctuationist, there is something very special about special stasis. Stasis, to him, is not just evolution that is so slow as to have a rate of zero. Stasis is not just a passive lack of evolution because there is no driving force in favor of change. Rather, stasis represents a positive resistance to evolutionary change. It is almost as though species are thought to take active steps not to evolve, in spite of driving forces in favor of evolution.”78
He also concedes that natural selection only serves to slow this rate of mutation, “Instead, what we find is that natural selection exerts a braking effort on evolution. The baseline rate of evolution, in the absence of natural selection, is the maximum possible rate. That is the mutation rate.”79
But if this theory were true, presumably scientists could test it by examining organisms that became geologically isolated in the fossil record. They would examine areas where glaciers had cut off an extinct population. Or, they would look at a population that became separated from a parent group on an island, like Hawaii or Madagascar. But Gould argued that most major evolutionary change takes place in isolated subpopulations so fast that there is insufficient time for the transitional forms to be fossilized in any place and at any time in history. Thus, the evidence that would support this theory miraculously disappeared in every time period throughout the history of the Earth and in every location across the globe where it happened “Speciation, the process of macroevolution, is a process of branching. And this branching … is so rapid in geological translation (thousands of years at most compared with millions for the duration of most fossil species) that its results should generally lie on a bedding plane, not through the thick sedimentary sequence of a long hillslope.”80
Dawkins translates Gould’s theory into plain English. He believes that the missing fossils will always be in places where we are not looking: “. . . most of the evolutionary change took place somewhere else! . . . most of the gradual change goes on in geographic areas away from the areas where most fossils are dug up.”81
In short, evolution cannot be refuted because the evidence that would refute it will always be hidden and beyond mankind’s ability to find it. Even though humans can find fossilized bacteria in Antarctica and Greenland, Gould and Dawkins believe that humans can never find the missing fossils in support of evolution.
But even if it were true that only the missing transitional organisms were beyond mankind’s ability to find, a theory that cannot be tested or falsified falls outside the definition of true science. According to the National Academy of Science, scientific theories must be based upon repeatable observations, subject to testing, and “falsifiable,” as observations could disconfirm the theory.82 During the 2005 trial related to intelligent design, the Court relied upon this definition of true science to test intelligent design: “As the National Academy of Sciences (hereinafter “NAS”) was recognized by experts for both parties as the “most prestigious” scientific association in this country, we will accordingly cite to its opinion where appropriate. (citation omitted.) NAS is in agreement that science is limited to empirical, observable and ultimately testable data: “Science is a particular way of knowing about the world. In science, explanations are restricted to those that can be inferred from the confirmable data—the results obtained through observations and experiments that can be substantiated by other scientists. Anything that can be observed or measured is amenable to scientific investigation. Explanations that cannot be based upon empirical evidence are not part of science.”83
The Court then found that intelligent design was not a science because the judge believed that it could not be properly tested: “Accordingly, the purported positive argument for ID [intelligent design] does not satisfy the ground rules of science which require testable hypotheses based upon natural explanations. ID is reliant upon forces acting outside of the natural world, forces that we cannot see, replicate, control, or test, which have produced changes in this world. While we take no position on whether such forces exist, they are simply not testable by scientific means and therefore cannot qualify as part of the scientific process or as a scientific theory.”84
If this is the correct standard for true science, it must also be applied to punctuated equilibrium. If punctuated equilibrium must be accepted as true because it is beyond any person’s ability to refute it, then it is not a scientific theory. Commentator Steven Stanley also calls this theory as unscientific: “Any claim that natural selection operated with great effect exactly where it was least likely to be documented -- in small, localized, transitory populations -- would seem to render Darwin’s new theory untestable against special creation, and perhaps almost preposterous as a scientific proposition.”85
(h) Punctuated Equilibrium’s Inability to Explain the Cambrian Explosion.
If we accept that punctuated equilibrium is a real scientific theory, can it explain the Cambrian explosion? Even Dawkins concedes that the answer is “no”: “My point here is that, when we are talking about gaps of this magnitude, there is no difference whatsoever in the interpretations of ‘punctuationists’ and ‘gradualists.’ Both schools of thought despise so-called scientific creationists equally, and both agree that the major gaps are real, that they are true imperfections in the fossil record. Both schools of thought agree that the only alternative explanation of the sudden appearance of so many complex animal types in the Cambrian era is divine creation…”86
Thus, if punctuated equilibrium cannot be tested and if it cannot explain the Cambrian explosion, it also holds no value in explaining other gaps in the fossil record. Scientific theories cannot be applied on an ad hoc basis.
(6) The Inability of Mutations to Explain the Cambrian Explosion.
Creationists agree that allopatric speciation and natural selection can create groups that have difficulty interbreeding. In these cases, creationists and evolutionists agree that the creation of a new species occurs from the loss of genetic information in the breeding pool. The new species in these cases are not the result of endless micro-mutations occurring at the cellular level where the DNA is copied or by some other means. Instead, creationists contend that change within an organism has boundaries. These boundaries of change show God’s intelligent design to allow species to survive changing environments.
For Darwin’s theory to be correct, evolutionists must identify a process whereby new information is added to allow for the creation of new species with entire body parts or new organs. The only process by which this change could theoretically occur is through the process of mutation, not natural selection. According to Dawkins, “Natural selection may only subtract, but mutation can add.”87
To explain how mutation cannot result in “macro-evolutionary” change, we consider: (1) the mutation rate claims made by advocates of punctuated equilibrium; (2) the lack of evidence for punctuated equilibrium at the molecular level; (3) the observed fatal or harmful results of most mutations; (4) alternative theories of helpful mutations lying dormant in a recessive state until a critical mass is reached; (5) problems with this alternative theory; (6) genetic research which contradicts the theory of macroevolution or limits it to viruses, bacteria, and possibly ants and termites; and (7) the inability of modern theories of mutation to explain the Cambrian explosion.
(a) Punctuated equilibrium’s 50,000-year speciation theory.
Gould and Eldredge claimed that punctuated equilibrium could allow species to form at incredibly fast rates. Gould argued that: “Punctuated equilibrium accepts the conventional idea that species form over hundreds or thousands of generations and through a series of intermediate stages.”88 He further argued: “I’d be happy to see speciation taking place over, say, 50,000 years…”89 Likewise, Eldredge estimates: “Geographic isolation leading to reproductive isolation need not take long to occur: our estimate was from five thousand to fifty thousand years.”90
Thus, under their theory, only when an organism is geographically separated and freed from natural selection are the breaks allegedly released and the evolutionary throttle allegedly kicks into high gear. Dawkins in fact suggests this to be the true normal gear. But he claims that the normal mutation rate for an organism is: “often less than one in one million.”91
(b) Punctuated equilibrium’s lack of evidence at the molecular level.
Although Gould and Eldredge’s theories of a variable genetic mutation rate are widely accepted amongst paleontologists, Dawkins concedes that most microbiologists who study change at the cellular level confirm that the rate of mutation is in fact constant, “Ironically, a form of constant speedism has recently become highly favored among modern molecular geneticists.”92
In order to allow for allegedly explosive periods of mutation when a subgroup is separated from the main group in the face of cellular evidence that the rate of change is constant, he suggests that there are in fact two rates of change at the cellular level. These include a constant rate that scientists can document and a separate faster rate that he believes to exists when a population is separated that cannot be documented: “evolutionary change at the level of the protein molecules really does plod along at a constant rate . . . even if externally visible characteristics like arms and legs are evolving in a highly punctuated manner.”93 Amazingly, Dawkins believes that protein molecules and physical characteristics evolve at the same rate until they are geographically isolated where they cannot be observed. At that point, for some unknown reason, the genetic mutation rates at the molecular and physical levels diverge.
Arguably, microbiologists who actually study change at the cellular level have more credibility to opine on the rate of change at the cellular level. Indeed, many microbiologists have been quick to point out there is no genetic evidence to punctuated equilibrium’s mutation theory: “[S]ome of the genetic mechanisms that have been proposed [by proponents of punctuated equilibrium] to explain the abrupt appearance and prolonged stasis of many species are conspicuously lacking in empirical support. Thus, we do not feel logically compelled to abandon neo-Darwinism in favor of the theory of punctuated equilibria.”94
(c) The frequently fatal result of mutations.
Observable genetic mutation is in fact neither a frequent nor a friendly event. The observed effects of mutation are almost always harmful to the individual organism. For this reason, microbiologists have historically believed that macro-evolution requires tremendous periods of time.
By studying the genes and rates of mutation, both good and bad, of the fruit fly Drosophila, the limits of mutation change at the genetic level are clearly established. As noted by one prominent biologist: “[Observable] mutations are not common. In fact, it has been calculated that a gene changes only once in every one hundred thousand to one million replications (reproductions). Another way to express the basic rate is to find out how many gametes contain at least one mutant gene. Studies show one mutant gene in ten to one hundred gametes.”95
Dawkins offers similar estimates when talking about what can be observed or documented: “In real life, the probability that a gene will mutate is often less than one in one million.”96 He adds that natural selection only serves to slow this rate of mutation.97 In the case of genes allegedly mutating to work together, he emphasizes that the process is particularly slow in that there are genetic forces which actively resist such change: “they resist invasion by new mutant genes which are not members of the club. This is quite a sophisticated idea, and it can be made to sound plausible.”98
Thus, changes on the genetic level are slow to occur. The chance of multiple observable genetic mutations are not only as unlikely as being struck by lightning, the effects are equally harmful. If the mutation rate were put on steroids when a sub-group became geographically separated in order to make the theory of punctuated equilibrium work, the organism would die. As one commentator explains: “In fact, only one in one thousand (mutations) is not harmful. Gene mutations can produce structural impairments and genetic diseases, to say nothing of when mutant genes are lethal. It is known that mutations produce harmful traits that natural selection actually selects against, or eliminates. Some lethal mutations, in fact, terminate the development of an individual as early as the zygote stage. Yet if macro-evolution by means of natural selection is to occur, there must first be new traits for which to select . . . most observed mutations are harmful, and there is no experimental evidence to show that a new animal, organism or even a novel structural feature has ever been produced from the raw material produced by mutations.”99
The genetic information necessary for life forms to function is much like the highly organized development of letters and words and sentences to form meaningful paragraphs. A copying error in an English paragraph will rarely make the paragraph clear. The analogy is the same in the biological world. However, where the English sentence merely becomes unclear through an error in copying, an error in copying in the genetic code of an organism frequently results in the death of that organism. We need only look to humans with genetic diseases to confirm this to be true.
On this point, Dawkins again agrees. He concedes that even small mutations are likely to be fatal, “the chances are very high that a big random jump in genetic space will end in death. Even a small random jump in genetic space is likely to end in death.” 100 He also concedes that there is no experimental evidence to show that a new animal, organism or even a novel structural feature (“macroevolution”) has ever been produced from the raw material produced by mutations. All Dawkins can argue in response is that “humans haven’t the patience to wait a million generations for a mutation.”101
In other words, if we are looking for some tangible proof of macroevolution, Dawkins claims that we do not find it in the area of observed genetic mutations. But if the theory cannot be refuted with observation or testing, it again falls outside the definition of true science.
(d) The theory of silent mutations laying hidden in a recessive state until a critical mass is reached.
Because punctuated equilibrium is not supportable at the molecular level, other evolutionists have searched for different theories that would allow for macroevolution to be plausible at the genetic level. But these theories all suffer from a similar flaw that they are neither based upon genetic research nor can they be tested or reproduced. For example, some modern advocates of macroevolution now contend that mutations might occur in regulatory genes that lie “silent” in a recessive state until a critical mass of mutations appear together for the changes to appear in the dominant state. Two leading evolutionary paleontologists, Ian Tattersall and Jeffrey Schwartz, cite this theory to explain how major changes appear “out of nowhere” in the fossil record. But they concede that they have no idea how or why this process might occur: “[W]hen these features appear, it will be as if ‘out of nowhere.’ They will also appear in their full-blown state in a number of individuals, because as recessives they will have remained ‘silent’ until a critical mass of them had accumulated in the population, permitting them to be expressed in the homozygous state.102 Finally, at some point, and by a mechanism that remains unknown, the mutant recessive is converted to the dominant state. Hence, then, we have a potential explanation at the genetic level for why, for instance, the acquisition of [the] modern human body form appears to have been virtually instantaneous, rather than the result of perfecting transformation over ages.”103
(e) Problems with the hidden mutations laying in a recessive state.
For at least three reasons, this alternative theory also fails.
First, there is no genetic evidence to support it. If, on the other hand, it is merely conjecture as to what might cause macroevolution, it is not proof that macroevolution in fact takes place.
Second, modern evolutionists concede that, if change is to occur, it must occur where members of a species are isolated from the others. As Dawkins explains, “groups of genes are so well adapted to each other that they resist invasion by new mutants which are not members of the club.”104 That is why we are told that changes do not continue to appear in humans. But where the members of the species are isolated, there can be no “critical mass” of silent friendly mutations waiting to be triggered. The problem of finding a critical mass is only compounded by the fact that “the probability that a gene will mutate is often less than one in one million.”105 Moreover, as previously shown, most mutations are not beneficial. They are instead either harmful or fatal.
Third, if a mutation did occur in a recessive state with no positive or negative effect, there is no reason why such a mutation would be passed on through multiple organisms to future generations. Sir Ronald Fisher, one of the world’s experts on the mathematics of evolution, showed that isolated changes are statistically unlikely to be passed down to future generations. “A mutation, even if favorable, will have only a very small chance of establishing itself in the species if it occurs once only.”106 Fisher’s calculations establish that, with only a single positive mutation, the odds are 500 to one against its survival. Based upon Fisher’s computations, biophysicist Lee Spetner estimates that there would have to be almost 350 mutants to have a 50 percent chance of survival. There would have to be 1,100 of them to have a 90 percent chance of survival.107 Using the 50 percent number as a benchmark, Tattersall’s and Schwartz’ theory becomes unworkable when we consider that the 350 “silent” mutations must accumulate in an isolated population with a mutation rate of less than one in one million, most of them harmful. Moreover, based upon the current theories of evolution, this must all happen in an isolated sub-group.
(f) Genetic research which contradicts or limits the theory of macroevolution.
Advocates of macroevolution can now have their theories tested against empirical research related to genetic mutations. Genetic laboratory research of the fruit fly with its rapid new generation rate in fact empirically refutes the mere possibility of macroevolution. It is now known that between thirty and forty genes operate in unity to code the wing structure of a fruit fly (see below) to allow the wing to exist and fly.108 Dawkins likewise concedes that large numbers of genes from different locations must work together to create new structures, “the whole process of embryonic development can be looked upon as a cooperative venture, jointly run by thousands of genes together. Embryos are put together by all the working genes in the developing organism in collaboration with one another.”109
E. J. Ambrose, Emeritus Professor of Cell Biology from the University of London, conducted studies related the number of genes needed to have the wing in the fruit fly, even at 1 percent of its size.
The minimum number of genes: He estimated that it “is most unlikely that fewer than five genes could ever be involved in the formation of even the simplest known structure previously unknown in the organism.” 110
The non-harmful mutation rate: He also found that: “It is a reasonable estimate that no more than one new non harmful mutation will occur per generation in a population of one thousand.”111
The probability of two non-harmful mutations: He further discovered that most genes have a mutation frequency smaller than one in one hundred thousand. The probability, then, of two non-harmful mutations occurring in the same organism is one in one million. (The probability of two independent events occurring is the product of their independent probabilities; thus one/one thousand x one/hundred thousand equals one/one million.) 112
The probability of five non-harmful mutations: The odds of five non harmful mutations occurring in the same individual are one in one thousand million million! (Figure this by multiplying the five independent probabilities: 10 -3 x 10 -3 x 10- 3 x 10 -3 x 10 -3.) For all practical purposes, there is no chance that these five mutations will all occur within the life cycle of a single organism, as any biologist would agree.113 The chance is even more amazing if we consider that between thirty and forty genes must mutate to provide us with even a minimally functioning fruit fly wing.
The problem of coordinating non-harmful mutations: Ambrose further found that the improbable coming together of five genes must further be joined with the random chance that all will be coordinated to work together. He notes that these problems: “fade into significance when we recognize that there must be a close integration of functions between the individual genes of the cluster, which must be integrated into the development of the entire organism.”114
Ambrose’s findings of serious problems with the theory of macroevolution: Based upon all these problems, Ambrose concluded: “We conclude therefore that recent hypotheses about the origin of species fall to the ground, unless it is accepted that an intensive input of new information is introduced at the time of isolation of the breeding pair.”115
Thus, the example of the fruit fly’s wing empirically undermines the possibility of genetic mutations leading to macro-evolutionary changes forming new organs, structures, or other new and distinct anatomical changes.
Ross points to other research which limits the possibility of positive mutations to bacteria, viruses and possibly ants and termites: “A fundamental problem biologists observe (for well understood biochemical reasons) is that deleterious mutations vastly outnumber beneficial manufactured mutations. Estimates range anywhere from a deleterious-to-beneficial ration of 10,000 to 1 up to 10,000,000 to 1. These numbers mean that only species with enormous populations, short generation times, and small body sizes would be able to survive long enough to advance through beneficial mutations. The onslaught of deleterious mutations coupled with environmental stresses – such as Earth’s declining rotation rate, the Sun’s increasing luminosity, changes in the chemical composition of the atmosphere, supernova eruptions, asteroid and comet collisions, solar flaring events, and climate cycles, among others - would drive most life-forms to extinction. Biologists directly observe significant evolutionary advancements only for those species exceeding a quadrillion individuals with body sizes less than one centimeter and generation times less than three months. To be specific, significant evolutionary advancements are confirmed by direct field observation for some viruses and bacteria species. It is debatable for ant and termite species. But that’s it.” 116
(g) The inability of genetic mutations to explain the Cambrian evolution.
Evolutionists respond to these problems with the theory of “exaptation.”117 This theory alleges that primitive organisms with parts with one function can be co-opted into a different function over time. Thus, they claim that multiple mutations all do not need to happen at the same time for a new structure to appear. But this theory has never been and cannot be tested with anything in the real world.
Even if their theory were valid, there were (with the one unrelated exception of the Ediacara biota) only single celled organisms prior to the Cambrian explosion. No theory of genetic mutation can explain the 70 to 75 phyla that miraculously appear within only five to ten million years.
For these reasons, even prominent researchers concede that modern theories of mutation cannot explain the Cambrian explosion: “It follows that 6-10 million years in the evolutionary time scale is but a blink of an eye. The Cambrian explosion denoting the almost simultaneous emergence of nearly all the extant phyla of the Kingdom Animalia within the time span of 6-10 million years can’t possibly be explained by mutational divergence of individual gene functions.”118
But if modern theories of mutation cannot explain the sudden appearance of all the 70 to 75 animal phyla that have ever existed, why should these theories be deemed true in explaining other gaps in the fossil record? Again, scientific theories are not scientific if they are only applied on an ad hoc basis.
(7) The Inability of Every Possible Predecessor Organism to Stay Hidden.
No matter what the facts may show, most evolutionists will never accept the possibility that God specially recreated a single organism in the history of the Earth, including during the Cambrian explosion. Indeed, the American Civil Liberties Union sees even the mere discussion about this possibility as a threat to both academic integrity and religious liberty: “To reject Creationism as science is to defend the most basic principles of academic integrity and religious liberty.”119
What then do evolutionists have to say when confronted with these multiple lines of evidence that there is simply no natural explanation for the Cambrian explosion? Their responses generally fall into one of two categories.
First, like Darwin, many will admit that they have no explanation and simply rely upon the promise that science will one day provide the answers. They claim that creationists are merely pointing to gaps in our scientific understanding to prove God’s existence. They call this the “God of the gaps fallacy.” In fact, the Cambrian explosion shows that evolutionists have a “Darwin in the gaps” fallacy. They have a faith that Darwin’s theories will one day explain the gaps in our understanding. After more than 150 years of searching for an answer, it’s safe to conclude that it has no natural answer to the Cambrian explosion. To continue to believe that science will one day provide an answer can only be described as a “faith” in naturalism.
Second, others argue that the alleged transitional organisms between the single celled protist and the invertebrates, the cordobites, boney lobed fish, insects, and others must have never fossilized. To support his theory, Dawkins points out that “many of these animals had only soft parts to their bodies: no shells or bones to fossilize.”120
But this defense does not withstand scrutiny. Just before the Cambrian explosion, rocks dating from 580 to 543 million years ago contain fossils of the “Ediacara biota”. These soft multi-celled organisms from the “Avalon explosion”, just before the Cambrian explosion, had no trouble being preserved in the fossil record.121 The pre-Cambrian fossil record also contains single-celled organisms dating back to shortly after the formation of the Earth. For Dawkins’ theory to be true, only the transitional organisms miraculously failed to fossilize.
Even Niles Eldredge, one of the founders of punctuated equilibrium, undermines this argument. He has pointed out that the fossil record from such remote places as Antarctica, Greenland and other areas is sufficiently complete for us know exactly when soft bodied bacteria and protists first formed: “Now we see an increasingly rich early fossil record - one restricted to bacteria for at least its first billion years - but rich enough for us to get some definite ideas of what was the earliest phases of life’s evolution were like.”122
The late Stephen Jay Gould also conceded that - -after more than 150 years of extensive searching - - the argument that the preCambrian transitional species could not fossilize is simply not true: “The most famous such burst, the Cambrian explosion, marks the inception of modern multicellular life. Within just a few million years, nearly every major kind of animal anatomy appears in the fossil record for the first time ... The Precambrian record is now sufficiently good that the old rationale about undiscovered sequences of smoothly transitional forms will no longer wash.”123
Commentators Ankerbert and Weldon point out that this pessimism is well founded. After more than 150 years of searching, scientists have amassed more than 250 million cataloged fossils.124 Ankerbert and Weldon quote a similar concession from a well-known geologist, David Rump from the University of Chicago: “‘We now have a quarter of a million fossil species but the situation has not changed much. The record of evolution is still surprisingly jerky and, ironically, we have even fewer examples of evolutionary transition than we did in Darwin’s time . . . ’”125
Speaking broadly and not limited to the Cambrian explosion, other paleontologists have conceded that the missing transitional species cannot be explained through an allegedly incomplete fossil record: “The record jumps, and all the evidence shows that the record is real: the gaps we see reflect real events in life's history -- not the artifact of a poor fossil record.”126 “Evolutionary biologists can no longer ignore the fossil record on the ground that it is imperfect.”127
In a moment of candor, Dawkins admits that his position regarding the inability of only the alleged pre-Cambrian transitional species to fossilize is in fact inconsistent with the extensive soft-tissue pre-Cambrian fossil record: “If you are a creationist you may think that this is special pleading.”128
By “special pleading,” Dawkins refers to inconsistent pleading. On this point, he is right. If macro-evolution is a scientific theory, it does not get to change the rules when the facts don’t support it.
Finally, evolutionists typically respond to these facts by claiming that creationists only make claims in the form of “negative argumentation” without making predictions that can be tested by science. But that is not true. It bears repeating that, thousands of years ago, Moses revealed that God created the animals of the sea in an instant of His time after He created plant life and before He created the land animals: “Then God said ‘let the waters team with swarms of living creatures, . . . (1:20). And God created the great sea monsters and every living creature that moves, with which the waters swarmed after their kind, . . .; and God saw that it was good (1:21). And God blessed them, saying ‘be fruitful and multiply, and fill the waters in the seas . . .’ (1:22). And there was evening and there was morning, a fifth day.” (1:23).
Moses’s amazing prediction of the life’s “Big Bang” in the oceans shows that the Bible is God’s inerrant word: “20 For since the creation of the world His invisible attributes, His eternal power and divine nature, have been clearly seen, being understood through what has been made, so that they are without excuse. 21 For even though they knew God, they did not honor Him as God or give thanks, but they became futile in their speculations, and their foolish heart was darkened. 22 Professing to be wise, they became fools, 23 and exchanged the glory of the incorruptible God for an image in the form of corruptible man and of birds and four-footed animals and crawling creatures.” (Ro. 1:20-23).
2. The First Amphibians: Evidence of God’s Intelligent Design. Gen. 1:20-23.
Amphibians and reptiles are not specifically listed after the fish in the Bible because God focused on the Kosher animals that humans could later eat. It is, however, possible that they are included with reference to the non-Kosher “swarming” creatures of the sea.
Even though amphibians are not specifically listed, they are addressed here because evolutionists claim that they formed the alleged transition from fish to land-based life. But amphibians also appear in the fossil record without transitional forms. Dr. Duane Gish quotes Robert Carroll, another prominent paleontologist, as conceding: “We have no intermediate fossils between Rhipidistian fish and early amphibians . . . “129
Approximately 370 million years ago during the Devonian period, the first amphibians appeared. Evolutionists now allege that they emerged from lobe-finned fish, which were similar to the modern “coelacanth” and “lungfish”.130 Evolutionists also allege that these ancient lobe-finned fish evolved into multi-jointed leg-like fins with digits that enabled them to crawl along the sea bottom. Evolutionists further allege that some fish had developed primitive lungs to help them breathe air when the stagnant pools of the Devonian swamps were low in oxygen. Under this evolutionary model, they could also allegedly use their strong fins to hoist themselves out of the water and onto dry land if circumstances so required. By processes that evolutionists can't explain, their bony fins then allegedly evolve into limbs. The missing transitional organisms then allegedly become the ancestors to all “tetrapods.” These are animals that include modern amphibians, reptiles, birds, and mammals. Under this evolutionary model, despite being able to crawl on land, many of the allegedly transitional fish still spent most of their time in the water. Evolutionists claim that they have started to develop lungs. But they still breathed predominantly with gills.131
Some paleontologists believe that a fish called crossopterygians (kraw-sahp-tul-rig-nz), a lobe-finned fish, was the first alleged transitional fish on the path to the first amphibians. A very similar lobe-finned fish swims in the Indian Ocean today. Some theorize a transition from lobe-finned fish to amphibians as depicted below:
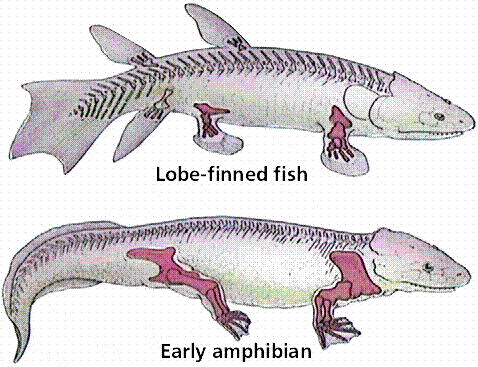
Source:132
But biologists Percival Davis and Dean Kenyon point out that the claim that crossepterygians was an alleged transitional species is based upon pure speculation: “If crossepterygians really did evolve into amphibians, tremendous changes must have taken place. Fins must have transformed into forelimbs. The skull had to change from two parts into a single, solid piece. The hip bones had to enlarge and become attached to the backbone. Numerous changes must have occurred in the organs, muscles and other soft tissues. For example, the air bladder of a fish had to be transformed into the lungs of the amphibian. Though just a few of the many examples possible, these are enough to show how large the differences between early fish and amphibians really were. How many different transitional species were required to bridge the gap between them; hundreds? even thousands? We don’t know, but we do know that no such transitional species have been found. Moreover, we have no fossil evidence of the evolution of the crossopterygians from other fish. Two large gaps thus exist in the fossil record between ordinary Devonian fish (325 million years ago) and amphibians; one between ordinary fish and crossopterygians, and an even larger gap between those lobe-finned fish and amphibians.”133
Because the amphibian and the fish have entirely different physiologies, different living environments, and other major differences, a transitional species would be readily identifiable if found. In summarizing all the evidence to date, Dr. Gish concluded before his death: “All the fish cited as being the most likely ancestors of amphibians are 100% fish which were required to spend all of their time in the water, while all of the so-called descendant amphibians were 100% amphibians with the basic amphibian limbs, feet, and legs. No one has succeeded in finding a single transitional form with part fins and part feet.”134
Dr. Gish further noted that there are thousands of different amphibian species, like salamanders, newts, frogs, and toads. After summarizing the writings of other paleontologists, he concludes: “When they first appeared in the fossil record, they were just as they are today, modern in appearance and diverse in structure.”135
Evolutionists, however, treat these questions with a collective yawn. Freed from the difficulty of answering questions about the Cambrian explosion, they claim that their theories of evolution can finally be applied. Under the theory of punctuated equilibrium, the transitional fossils are simply hidden in a place where scientists cannot find them. Furthermore, the theory of “exaptation” allegedly explains how primitive organisms with parts with one function can be co-opted into a different function over time. Even though these theories cannot be tested against anything in the fossil record, the theories are enough for evolutionists to satisfy themselves that they have explained how fish evolved into amphibians.
But in addition to failing the test of being a reputable scientific theory, these theories also fail when the alleged transition between fish and amphibians is examined at the molecular level. A common protein found in all living organisms is called “cytochrome C”. Evolutionists claim that cytochrome C provides a molecular clock against which evolution can be tested. For example, Dawkins asserts: “[W]e can measure exactly how many steps separate one animal from another, at least with respect to a particular protein molecule . . .any one kind of molecule seems to evolve at a roughly constant rate in widely different animal groups. This means that the number of differences between comparable molecules in two animals, say between human cytochrome and warthog cytochrome is a good measure of the time that has elapsed since their common ancestor lived. We have a pretty accurate ‘molecular clock.’ The molecular clock allows us to estimate, not just which pairs of animals have the most recent common ancestors, but also approximately when these common ancestors lived.” (emphasis in original).”136
The National Academy of Sciences also cites to this alleged protein clock to attack the science of irreducible complexity: “[S]tructures and processes that are claimed to be “irreducibly” complex typically are not on closer inspection. For example, it is incorrect to assume that a complex structure or biochemical process can function only if all its components are present and functioning as we see them today. Complex biochemical systems can be built up from simpler systems through natural selection. Thus, the “history” of a protein can be traced through simpler organisms …” 137
But when the cytochrome C protein test is applied to fish and amphibians, the test reveals that fish did not evolve into amphibians. Dr. Michael Denton, a British –Australian biochemist, explains that the carp, a type of fish, has a thirteen percent difference in its molecular arrangement of the protein cytochrome C from the next alleged evolutionary species, the amphibian. He points out that, if the theory of macro-evolution were true, the molecular difference between the carp and mammals, like the horse or the rabbit, should be greater than the molecular difference between the carp and an amphibian, like the bullfrog. In fact, they are exactly the same. He concludes: “Again, an extraordinary mathematical exactness [thirteen percent] in the degree of isolation is apparent. So, although cytochrome C sequences varied among the different terrestrial vertebrates, all of them are equidistant from those of fish. At a molecular level, there is no trace of the evolutionary transition from fish to amphibian to reptile to mammal. So, amphibia, always traditionally considered intermediate between fish and other terrestrial vertebrates, are in molecular terms as far from fish as any group of reptiles or mammals! To those well acquainted with the traditional picture of vertebrae evolution the result is truly astonishing.”138
Dr. Denton further notes that, on a molecular level, some amphibians are as close to other mammals as other amphibians: “For example, the degree of molecular divergence among frogs, which are morphologically very similar, is as great as that between mammals, which are morphologically very diverse. . . . There is little doubt that if this molecular evidence had been available one century ago it would have been seized upon with devastating effect by the opponents of evolutionary theory like Agassiv and Owen, and the idea of organic evolution might never have been accepted.”139
Thus, applying the same molecular test that allegedly proves evolution between other organisms, fish did not evolve into amphibians. Instead, like fish, amphibians appear out of nowhere in the fossil record with no natural explanation that can explain their origin.
3. Reptiles: Evidence of Intelligent Design.
Reptiles were certainly known to Moses. But he also did not list them because they are not Koshier animals. Nevertheless, they are discussed here because they represent the next alleged evolutionary step on the pathway to humans.
Approximately 310 to 320 million years ago during the late Carboniferous period, reptiles first appeared.140 Some claim that the oldest known reptile was an animal now called a “Casineria.”141 Others claim that the oldest confirmed reptile lived in Nova Scotia Canada approximately 315 million years ago. A series of footprints allegedly show reptilian toes and imprints of scales. The reptile is now called “Hylonomus.”142 It was a small, lizard-like animal. It was approximately about 20 to 30 centimeters (or 7.9 to 11.8 in) long. It also had numerous sharp teeth and most likely lived off insects.143
Like the alleged transition from fish to amphibians, the alleged transition from amphibians to reptiles is unsupported by transitional species and equally mysterious to evolutionists. Moreover, like amphibians, reptiles diverge widely to include flying reptiles, marine reptiles, gliding reptiles, snakes, turtles, and dinosaurs. For each class of reptiles, evolutionists have failed to offer any transitional species.
Robert L. Carroll is widely considered to be an authority on vertebrate paleontology. In 1988, he conceded that no known transitional organisms exist establishing an evolutionary link between amphibians and reptiles: “The early amniotes [animals that lay eggs like reptiles] are sufficiently distinct from all Paleozoic amphibians that their specific ancestry has not been established.”144
In 1997, he again conceded that the evolutionary origin of the reptile remained unknown: “The origin of the modern amphibian orders, (and) the transition between early tetrapods [land based animals including reptiles] are ‘still poorly known’.”145
For a brief time, evolutionists held up an organism named “seymouria” as a possible transitional link between amphibians and reptiles. However, Stephen Jay Gould later established that this could not have been a transitional species because reptiles predate the appearance of this organism by at least 30 million years.146
The absence of any transitional species, however, poses no dilemma to the evolutionist. Under the theory of punctuated equilibrium, evolutionists believe that transitional fossils are simply hidden in places where scientists cannot find them. Furthermore, no matter how different amphibians are from reptiles, their theory of “exaptation” states that amphibians must have developed special features that were later co-opted into a different function to allow an organism to become a reptile. In other words, the evolutionists believe that there must have been creatures that were half amphibian and half reptile until they evolved into reptiles. The absence of any evidence to support that this in fact happened does not matter. For most, the mere theory that it could have happened is enough to establish that it did happen.
But a theory that transitional organisms are hidden is not a scientific proposition because it cannot be tested or refuted. Moreover, the alleged transition between amphibian eggs to reptilian eggs provides an opportunity to test the theory of exaptation. In other words, is it possible to have an organism that is “half amphibian” and “half reptile”? For several reasons, the answer is “no.”
The reptilian egg is radically different from the amphibian egg. Amphibians lay their eggs in water. Their eggs are transparent and have permeable membranes. These eggs possess an ideal structure for development in water.

Frogs with anamniotic eggs:147
By contrast, reptiles lay hard-shelled eggs, also called an “amniotic egg.” The amniotic egg allows air in. But it is impermeable to water. While an amphibian egg can obtain water and some of the things it needs from the surrounding water, the reptilian egg is like a space capsule. It allows the reptile to survive in a foreign dry environment. The egg must be hard enough to survive on the land. But at the same time the embryo must have a sharp tooth or caruncle - -things that do not exist with amphibians - - to break free from the egg.

Young crocodile emerging from its egg148
Dr. Denton explains how the transition between these two types of eggs cannot happen in partial steps. Without at least eight components appearing together, the reptilian embryo would die: “The evolution of the amniotic egg is baffling. It was this decisive innovation which permitted for the first time genuinely terrestrial vertebrate life, freeing it from the necessity of embryological development in an aquatic environment. All-together, at least eight quite different innovations were combined to make the amniotic revolution possible: [1] the formation of a tough impervious shell; [2] the formation of the gelatinous egg white (albumen) and the secretion of a special acid to yield its water; [3] the excretion of nitrogenous waste in the form of water insoluble uric acid; [4] the formation of the amniotic cavity in which the embryo floats (this is surrounded by the amniotic membrane which is formed by an outgrowth of a mesodermal tissue. Neither the amniotic cavity nor the membrane which surrounds it has any homologue in the amphibian); [5] The formation of the allantois from the future floor of the hind gut as a container for waste products and later to serve the function of a respiratory organ; [6] the development of a tooth or caruncle which the developed embryo can utilize to break out of the egg; [7] a quantity of yolk sufficient for the needs of the embryo until hatching; and [8] changes in the urogenital system of the female permitting fertilization of the egg before the hardening the shell.”149
Theory of “exaptation” imagines that some transitional organisms had some of the component parts being used in an amphibian for something else. The component parts were then slowly co-opted into the new use as a reptile. Thus, under this theory, some amphibians in the past developed a tooth or caruncle as a result of one set of mutations. When some mutant amphibian egg became hardened in one its descendants, the mutant amphibian had no problem breaking free from the hard egg. Thus, under this theory, the first reptile appeared.
The absence of any evidence of amphibians with a tooth or caruncle in the fossil record does not matter. The theory that one could exist is enough to establish that it in fact existed.
But if there were a “half-amphibian” “half-reptile” organism that only had a few of these eight things working together, the organism would die. It would be like sending an astronaut in a space capsule and having to choose between food, water and air. Without everything working together, the embryo dies.
Or, if the theorized transitional organism had the hard shell without a tooth or caruncle, the organism would again die without being able to break free. It would be like giving the astronaut no way to escape from the capsule. He or she would live only until his or her food, air and water ran out.
Or, if the organism had the tooth or caruncle without the hard shell, the organism would break the membrane of the egg sack prematurely and die. It would be like sending the astronaut into space with a thin plastic hull instead of steel.
We can also test the possibility of all eight mutations happening together without killing the organism. Using the probability theories developed by E. J. Ambrose, Emeritus Professor of Cell Biology at the University of London, the chance probability of these eight changes simultaneously occurring in the reproductive system can be easily measured. As described in the beginning of this study, no more than one new observable, non-harmful mutation will occur per generation in a population of one thousand. Most genes have a mutation frequency smaller than one in one hundred thousand. The probability of two observable, non-harmful mutations occurring in the same organism is one in one million (1/1000 x 1/1000 = 1/1,000,000.) The odds of five observable, non-harmful mutations is the equivalent of one thousand million million. Here, to have the eight observables, mutations occur simultaneously in order for the reptilian egg to survive is a chance event of 1 in 1023. In standard notation, this probability is the equivalent of one followed by twenty-four zeros.150
The number 1023 is a probability event that few could imagine. To put this number in perspective, it is estimated that there are only 1022 planets within the entire universe. 1022 is the equivalent of 10 billion trillion planets. Accordingly, the probability of obtaining a reptilian egg with all of the interrelated requirements for survival necessary is a number greater than all the planets in the universe!
Thus, the theory of “exaptation” cannot explain the transition from the amphibian to the reptilian egg. Any small change in the development of the egg would most likely be fatal to the developing embryo.
If the theory of exaptation doesn’t work for the first reptilian egg, it cannot be applied as a theory elsewhere in the fossil record. Scientific theories cannot be applied on an ad hoc basis. For these reasons, the reptilian egg is evidence of irreducible complexity and God’s intelligent design.
4. Winged Animals: The Bible’s Accurate Description of their Order of Appearance. Gen. 1:20-23.
Winged animals are not part of the alleged evolutionary line leading from bacteria to mankind. But they are included here for two reasons. First, Moses lists them in the Genesis account. Second, evolutionists consider the appearance of the bird to be their best evidence of macroevolution.
First, Moses offers another test for scientists to confirm the accuracy of the Bible. He lists the winged creatures as appearing after the first fish and other sea creatures, and before the appearance of the first land mammals (Gen. 1:20-24). Until the 19th Century, believers had only their faith to defend this claim. But discoveries between the 19th Century and the present have confirmed the accuracy of Moses’ claims. Moses would have no way of independently guessing whether birds appeared before or after land mammals. Many creation accounts in fact have all the animals appearing at the same time.
Approximately 235 million years ago, the first known winged creature appeared. It was called the “kuehneosaurs”. It lived in the Late Triassic period and was found in southwest England. It measured 72 centimeters long (2.3 feet), and it had “wings” formed from ribs which jutted out from its body by as much as 14.3 cm.151 But it could not fly. It could only glide.
Winged animals continued to appear while dinosaurs lived on the earth. Approximately 228 to 66 million years ago during the late Triassic period until the end of the Cretaceous period, pterosaurs roamed the skies. Evolutionists allege that the pterosaurs, the pteranodon and other avian dinosaurs were in fact types of birds. This is based upon their nesting behavior, their eggs, their other behaviors, their physiology, and the fact that both had feathers. This conclusion is disputed. But if this is true, these avian dinosaurs again confirm the Genesis ordering to be accurate.
Birds also deserve mention here because advocates of macro-evolution point to the origin of birds as allegedly offering the best evidence and one of the few alleged transitional fossils in support of their theory. In 1860, only a year after Darwin published “On the Origin of Species” a fossilized feather was found in a limestone deposit dated to the late Jurassic period. The following year, Christian Erich Hermann von Meyer described this feather as “Archaeopteryx lithographica.”152 This organism lived 150 million years ago in modern day Germany and most likely other parts of Europe.
In 1863, scientists put together a nearly complete skeleton for this animal. They described many features that made it appear to look like a reptile, including clawed forelimbs and a long, bony tail.153 Thomas Huxley, one of Darwin’s biggest advocates, called “Archaeopteryx” (see below) the transitional fossil between birds and reptiles. He also saw this as the proof of Darwin’s theory.154

Source:155
But other advocates of evolution, like Harry Seeley, argued that the similarities between “Archaeopteryx” and dinosaurs were due to “convergent evolution”.156 Convergent evolution is the belief that the evolution of different lineages will sometimes create analogous structures that have similar form or function. Their analogous structures, however, were not present in the last common ancestor for each groups 157 Thus, analogous structures can offer misleading evidence of evolution.
For example, the pterosaurs (1) (see below), the bat (2) (see below) and the modern bird (3) (see below) all have analogous wings. But they do not share an allegedly close common ancestor. They also have very different alleged evolutionary trees. Thus, their similar wings are not proof that one evolved from the other.

Source:158
For these reasons, similar structures are not alone proof of evolution. More importantly, new research establishes that Archaeopteryx was simply one of many garden variety feathered dinosaurs. New research also establishes a number of significant differences between Archaeopteryx and modern birds.
Most importantly, Archaeopteryx had a completely different skull from modern birds. Like a dinosaur, it had a true set of jaws that were equipped with many small, sharp teeth. By contrast, birds have beaks. Like dinosaurs, Archaeopteryx had a long, stiff, bony tail. By contrast, this feature is not present in modern birds. Like some winged dinosaurs, each Archaeopteryx wing had two separate fingers on the leading wing edges. These fingers had sharp, curved claws. By contrast, this feature is also not present in modern birds. Paleontologists are not even certain that it could fly.
Discoveries in the Liaoning Province of northeast China further demonstrate that many dinosaurs had feathers. These dinosaurs include: “Sinosauropteryx” and “Sinornithosaurus”.159 Likewise, other things like the wishbone, that once distinguished Archaeopteryx from dinosaurs, have now also been found in dinosaurs. For these and other reasons, paleontologist Lawrence Witmer at Ohio University asserts: “We've started finding a lot of dinosaurs with feathers, and lots and lots have a wishbone, even T. rex, for this progressive erosion of the avian status of Archaeopteryx.”160
Paleontologist Xing Xu at the Chinese Academy of Sciences in Beijing has likewise concluded that neither Archaeopteryx nor a similar creature in China “Xiaotingia” belong in the avian family. Instead, they are just two of many similar types of dinosaurs: “When the researchers analyzed features of Xiaotingia and Archaeopteryx, the resulting family tree clustered them together. Unexpectedly, it also yanked them out of the avialan category and placed the duo with the deinonychosaurs.”161
Even those who claim that the path of evolution between dinosaurs to birds is undisputed concede that the fossils proving this fact are missing. For example, in the book “Why Evolution is True”, author Jerry Coyne concedes: “We can certainly trace the evolution of birds from feathered reptiles, but we’re not sure exactly which fossil species were the direct ancestors of modern birds.”162
Finally, on June 23, 2000 in the journal of Science, scientists classified Longisquama insignis. It had feathers and appeared approximately 240 million years ago in Central Asia, before most avian dinosaurs first appeared. With this new discovery, Ornithologist Alan Feduccia at the University of North Carolina, a coauthor of the new study, called the theory that birds are related to dinosaurs “a delusional fantasy.”163 Even the Smithsonian Institute has publicly endorsed the interpretation that Longisquama insignis had feathers and existed more than 220 million years ago.164
For all these reasons, Archaeopteryx is not a transitional species. Thus, one of the few alleged transitional species that allegedly proved macro-evolution to be true has disappeared. Therefore, while the flying animals offer no support for the theory of evolution, they confirm Genesis to be accurate.
5. Mammals: Evidence of God’s Intelligent Design. Gen. 1:24-25.
In the Genesis account, Moses made another bold revelation. Thousands of years ago, he asserted that the first land mammals came after the fish and winged animals, yet before humans. (Gen. 1:24-25). His two-verse listing of the mammals obviously does not list every kind of the thousands of mammals that have existed. He instead focuses on the mammals (both Kosher and non-Koshier) that God created to help mankind.
Moses again did not have access to the fossil record. If he were copying off other creation accounts, he would have come up with a different order. Or, he would have claimed that all the mammals appeared together with other mammals.
For thousands of years, believers had only their faith to defend Moses’ claims. But beginning in the 19th Century and continuing through the present, scientists have again confirmed that Moses’ ordering is correct. This again reveals that the Bible was not merely penned by mankind. It was instead inspired by God.
God’s creation of the mammals is confirmed by three things. These include: (1) the “Big Bang” of mammal life without transitional forms; (2) the weak evidence of the alleged transitional animals between reptiles and mammals; and (3) the irreducible complexity and vast differences between mammals and other forms of life.
(1) The Big Bang of Mammal Life without any Transitional Forms.
In another explosion of life on Earth, God filled the planet with most mammals in a geologically short period of time. Before considering the timing of the appearance of most mammals, consider how diverse mammals are as a grouping.
The class “Mammalia” comprises a diverse group of animals with 32 different “orders” or sub groups. All mammals are warm-blooded. Most female mammals have mammary glands that produce milk for their young. Most mammals are placental mammals. This means that they give birth to their young.
“Eutherian” mammals, like primates, give birth to well-developed young. By contrast, “Marsupialia” mammals give birth to young animals that continue to develop in the mother’s pouch. These include kangaroos, wallabies, the koala, possums, opossums, wombats, and the Tasmanian devil.
A third group, called “monotreme” mammals, lay eggs. Examples include the duck billed platypus, the echidna, and the spiny ant-eater. In addition to laying eggs, the monotremes differ from other mammals in that they have lower body temperatures. They also have different legs which extend out and then vertically below them. While some of these like the duck-billed platypus might look like a descendant of birds, evolutionists do not claim that they shared an immediate common ancestor. Instead, evolutionists claim that their similarity is an example of “convergent evolution”.
Although milk production is common to all mammals, there are also significant differences in how they deliver milk to their young. With most mammals, the mother nurses her young through a nipple. But the duck-billed platypus releases milk through ducts in its abdomen. Likewise, in the case of one type of mammal, the Dayak fruit bat, the male produces the milk production for the newborn bat.
Bats are also the only mammal that flies. They also use an echo location system that is not similar to any land animal. Thus, for evolution to be true, we should expect to find multiple kinds of transitional species to each of these very different kinds of mammals.
The first appearance of most mammals presents another dilemma for the theory of evolution. Like everywhere else in the fossil record, mammals abruptly appear and without any transitional species. Moreover, as shown in the chart below, these extremely different kinds of mammals all appear at virtually the same time in the geological record in another “big bang” of life.
Source: 165
Modern discoveries have revealed only a few corrections to the chart above. The oldest confirmed monotreme or egg-laying mammal is the “Castorocauda”. It was a furry beaver-like animal with webbed feet. It lived approximately 164 million years ago in China.166 It has no known transitional species before it. The oldest confirmed fossil of placental mammals is the “Eomaia scansoria.” It lived 125 million years ago in China. It was a shrew-like species that was only five inches (14 centimeters) long and weighed no more than 0.9 ounces (25 grams).167 It also has no known transitional species before it. As shown on the chart above, neither had an immediate common ancestor because their bodies are radically different.
Even with these two exceptions and one group discussed below, vertebrate paleontologists call this a mammal “explosion” in the fossil record because of the absence of transitional forms leading to their appearance: “There are all sorts of gaps: absence of gradationally intermediate 'transitional' forms between species, but also between larger groups -- between, say, families of carnivores, or the orders of mammals.”168
Other studies have likewise labeled the appearance of most mammals in the fossil record as an “explosion” due to the absence of transitional forms and their sudden appearance.169 Even the most common and allegedly unsophisticated mammal, the rat, is without any known preceding transitional species. Romer concedes: “The origin of the rodents is obscure. When they first appear in the late Paleocene, in the genus paramys, we are already dealing with a typical, if rather primitive, true rodent, with the definitive ordinal characters well developed. Presumably, of course, they had arisen from some basal insectivorous, placental stock, but no transitional forms are known.”170
If evolution were true, we would not expect an “explosion” of extremely diverse kinds of mammals at the same time. We would also expect to find a long transition of fossilized transitional species leading to each of these very different kinds of mammals found today. But after more than 150 years of searching, that fossil evidence for the vast majority of mammals simply does not exist.
(2) The Weak Evidence of an Alleged Evolution from Reptiles and Mammals.
Of all the thousands of mammals, evolutionists only claim to have a few allegedly transitional fossil organisms. The most discussed mammals with alleged transitional species in the fossil record include: (1) double jointed jawed mammals; (2) hominids; (3) the whale; and (4) the horse. The whale and the horse trees have been disputed by others. But they are beyond the scope of this study. The allegedly double jointed jawed mammals are discussed here because they form the alleged link between mammals and reptiles. Hominids are discussed in the next section.
The oldest mammal with an allegedly double jointed jaw is the Morganucodontidae. It allegedly coexisted with the dinosaurs in the late Triassic to early Jurassic periods, almost 200 million years ago. Fossils from this group have been discovered in Europe, Southern Africa, and Eastern Asia. 171 The second group is the family Haramiyidae. It appeared at approximately the same time as the family Morganucodontidae. Fossil remains from this animal, however, have only been found in Whales, England, and Germany.172 The third group is called the Multituberculata. It appeared in the late Jurassic and early Cretaceous Periods, approximately 142 million years ago. They have been discovered in the deserts of Mongolia.173
Many evolutionists allege that the families Morganucodontidae and Haramiyidae represent an evolutionary link between reptiles and mammals because of their jaws. In short, many paleontologists claim (based upon disputed jaw reconstructions) that they had double joints in their jaws. By contrast, modern mammals do not: “Morganucodonts were equipped with an unusual structure, the ‘double-joint’, which meant that the jaw articulation would be made up of the dentary-squamosal joint as well as a quadrate-articular one. Both the articular and quadrate would eventually become the malleus and incus. There is a trough at the back of the jaw that houses post dentary bones, such bones are absent today in mammals . . ..”174
As a preliminary matter, this hardly seems like exciting proof of evolution. If correct, this only establishes that these mammals had two joints in their jaws. With 32 diverse orders of mammals in the land, sea and air, a double-jointed jaw hardly seems surprising.
More importantly, there is evidence that this alleged double joint is a mere misconstruction of the fragmented fossils available. Not a single intact lower jaw is available from either such mammal. All such specimens have been reconstructed from scattered fragments.175
Questioning the placement of this allegedly double jaw-joint, Dr. Gish concluded that the unique physiology behind each type of jaw can hardly be imagined to support two entirely different types of physical requirements: “Whether the denture bone of these creatures actually made contact with the squamosal can only be inferred. If there had been such a contact between the denture and the squamosal, could it be said that this constituted a mammalian jaw-joint which existed along-side the reptilian jaw-joint? We must remember that these creatures had a fully-developed powerful reptilian jaw-joint. The anatomy required for such a jaw-joint, including the arrangement and mode of attachment of musculature, the arrangement and location of blood vessels and nerves, etc., must be quite different from that required for a mammalian jaw-joint. How could a powerful, fully functional reptilian jaw-joint be accommodated along with a mammalian jaw-joint?”176
Dr. Gish further found that many of the same paleontologists who uphold these animals as allegedly transitional disagree as to whether the jaw-joints have been properly reconstructed. He quotes Kemp, the Author of Mammal-Like Reptiles as conceding: “in fact, there is some doubt whether there is actual contact between these bones.”177 Likewise, he quotes another prominent paleontologist, C.E. Gow, Paleontologia Africana, 24:15 (1981) as conceding, “more specifically, it is thought to have both [a] reptilian and mammalian jaw-joint. However, several of Crompton’s interpretations of the morphology of the lower jaw and its articulation with the skull were wrong; some but not all of these he has conceded to in print.”178
The different location of the jaw joints in reptiles and mammals also correspond with different bone structures which allow the animal to hear. Dr. Gish describes in technical detail the changes required to allow the jaw structure to change between a reptile and mammal while the animal continues to be able to hear: “[E]volutionists believe that as the bones of the reptilian jaw, except for the dentary, gradually became relieved of their function in the jaw, they were now free to either evolve out of existence or to assume new function. Thus, the quadrate and articular bones became free (they were by the way, firmly attached to the dentary in morganucodon) and somehow worked their way into the middle ear to eventually become the incus and malleus, respectively. This would have required that the stapes (columella) of the reptile became free from its attachment to the tympanum (eardrum) and the retro-articular process of the articular gained an attachment to the tympanum (since the articular bone of the reptile supposedly became the malleus of the mammal, which is attached to the tympanum). Somehow while all this was going on, the quadrate bone of a reptilian ancestor must gain its freedom, move into the middle ear, and insert itself between the stapes and malleus. While all this maneuvering was going on, all of these bones must have somehow been refashioned and re-engineered in the most miraculous manner so that they could function in an entirely new auditory apparatus.”179
As complicated and as hard to imagine as the change in bone structure while allowing the organism to hear while a change in the jaw structure occurs, the creation of an entirely new hearing organ in mammals, called the Corti, is even more difficult to explain or imagine. The Corti allows mammals to hear specialized sounds which reptiles are unable to detect. This highly complex organ is associated with the molecular pathways within the brain allowing such sounds to be translated and understood by the brain. While the jaw structure was allegedly re-engineered through evolutionary changes, this organ appeared de novo. Likewise, these changes occurred simultaneously with all of the required changes in the internal organ structure. For all these reasons, the allegedly double-jointed jaws in these organisms do not establish that they evolved from reptiles.
(3) The Irreducible Complexity and Vast Differences between Mammals and Reptiles.
None of these three groups of animals can be linked to reptiles through any transitional species. More importantly, the alleged transition between cold-blooded reptiles and warm-blooded mammals with entirely different internal organs and body systems has posed a challenge that no evolutionary theorist has yet answered.
According Dr. Michael Denton, multiple highly complex organisms must all simultaneously appear within the first mammal to avoid sudden death. These include: (1) the ability to regulate body temperature through hair, sweat glands, and specialized skin blood vessels; (2) increased mobility in order to obtain an estimated 10-fold increase in food to maintain the higher cellular metabolic rate; (3) a specialized diaphragm, where none previously existed, to allow for a greater rate of exchange of external gasses to sustain the higher metabolic rate; (4) a specialized heart and circulatory system to maintain a higher blood pressure; (5) specialized kidneys capable of filtering the blood and manufacturing hypertonic urine, each capable of excreting noxious waste products without dehydrating the animal; (6) a highly complex endocrine system to determine the appropriate rate of secretion and reabsorption in the kidney tubes; (7) specialized muscles capable of moving the organism at faster rates and sustaining movement during high aerobic rates of activity; (8) a higher developed sensory and central nervous system, necessary to maintain the balance between these various organisms; (9) A highly complex blood-clotting system; (10) specialized teeth including incisors, canines, and molars, necessary for the increased and specialized feeding requirements; (11) specialized jaws, capable of moving sideways and front to back or in circular movements to allow for the quicker swallowing of food; (12) specialized digestive systems capable of extracting additional and specialized nutrients for the warm-blooded mammal; (13) specialized mammary glands in females, necessary to produce milk for the young; (14) a specialized scrotum for the males, capable of regulating the temperature of the male sperm by bringing the testicles closer to or further away from the body as the external temperature changes; and (15) countless other specialized organs.180
One prominent evolutionary paleontologist, T.S. Kemp in his book Mammal-like Reptiles and the Origin of Mammals (New York Academic Press, 1982) p. 312-313, estimates conservatively that as many as 20-structures and processes of the mammal are necessary to maintain a constant internal environment to withstand external fluctuations. Commenting upon this “integrated homoeostatic mechanism,” Kemp concludes that each of these processes must have simultaneously appeared in the first mammal organism, or the organism would have died: “To conclude, the essence of mammalian biology is the very high degree of complexity and internal integration of the various structures and functional processes . . . evolution toward mammalian levels of homoeostasis involved practically all aspects of the organism simultaneously. No single structure or function could evolve very far without being accompanied by appropriate changes in all other features.”181
Dr. Denton further notes that the brain structure of the mammals is distinct from all other organisms: “Another diagnostic characteristic of mammals is the cerebral cortex. This is a complex outgrowth of neural tissue which forms the outer layer of the brain and which is the seat of all the higher mental functions and complex behavior patterns so characteristic of mammals. It consists of millions of nerve cells and gives rise to a number of neural pathways which are quite unique to the mammalian brain. In all mammals the nerve cells in the cortex are organized into the same invariant pattern of six basic layers of cells and the same regions are devoted to the analysis of the same sorts of information, visual, motor or sensory. Although the cortex varies enormously in different mammalian groups, being highly developed in primates and the Cetacea (the whales and dolphins), but only poorly developed in insectivores (the shrews and hedge hawks), its basic design is fundamentally invariant. There is nothing like it in the brain of any non-mammalian vertebrate and it is not let up gradually through a sequence of less complex neurological structures in any known group of organisms.”182
In addition to the required simultaneous appearance of so many irreducibly complex organs, the limbs of the mammals develop from entirely different parts of the vertebrae than any other vertebrate organisms. Dr. Denton refers to this evidence to question whether mammals belong in the same family group with fish, amphibians, and reptiles: “Another class of organs considered to be distinctly homologous [similar in appearance] are the vertebrate forelimbs, yet they generally develop from different body parts in different vertebrate species. The forelimbs develop in the trunk segments [the number of the vertebrae] two, three, four, and five in the newt, segment six, seven, eight, and nine in the lizard, and from thirteen, fourteen, fifteen, sixteen, seventeen, and eighteen in man. It might be argued that they are not strictly homologous at all!”183
Evolutionists offer no explanation for how so many, highly developed, complex, interrelated organisms could appear simultaneously. Indeed, the theory of gradual mutations contradicts such a proposition entirely. Evolutionists also cannot explain how these different structures could have existed in reptiles without killing the reptile and then becoming co-opted into the first mammal.
As previously stated, changes of any large nature are almost always known to be fatal to the organism. The development of even one such new organ would represent a substantial leap from the predecessor reptile organism. No evolutionary theory has or could explain such a massive change of so many new internal organs, simultaneously.
Proof of God’s intelligent design of mammals is also evident through the various types of highly specialized mammals, all appearing without transitional species. Dr. Gish provides the examples of the echolocation system of bats as an example of where evolutionary theory fails to provide any explanation. Bats are the only flying mammals and are considered to be some of the most specialized of all mammals known to exist. Bats emit a stream of high-pitched sounds which bounce off objects. The echo returns and is received by the very sensitive auditory apparatus of the bat. Millions of electronic impulses are transmitted to the brain each second, and the brain converts these electronic messages into an image. While thousands of bats may emit such signals together, each bat is capable of recognizing its own signal as being distinct from all others. Dr. Gish notes that when bats first appeared, approximately 50 million years ago, records reveal that they were virtually indistinguishable from modern bats. In short, bats with their highly developed echo-location and flight systems appeared without any trace of transitional species.
The proof of God’s intelligent design is not limited to such highly complex mammals as the bat. Even more iconic mammals, like the giraffe, are irreducibly complex in their body anatomy and cannot be explained under a theory of gradual mutation or co-opted change. Evolutionists have long hypothesized that the giraffe might have evolved from a horse-like creature. Evolutionists had speculated that the neck of this ancestral horse-like creature might have evolved to grow longer and longer into the present form of the giraffe. Given that the giraffe can eat leaves and other high up vegetation that other land mammals cannot, evolutionists have speculated that the giraffe provides a successful example of natural selection at work.
As scientists have gained greater understanding of the anatomy of the giraffe, this theory of gradual mutation becomes inadequate to explain the appearance of the giraffe. Biologists Percival Davis and Dean Kenyon provided the following description of the complex circulatory system of the giraffe: “The giraffe requires a very special circulatory system. When standing upright, its blood pressure must be extremely high to force blood up its long neck; this, in turn, requires a very strong heart. But when the giraffe lowers its head to eat or drink, the blood rushes down and could produce such high pressure in the head that the blood vessels would burst. To counter this effect, the giraffe is equipped with a coordinated system of blood pressure controls. Pressure sensors along the neck’s arteries monitor the blood pressure and activate contraction in the artery walls (along with other mechanisms) to counter the increase in pressure. In short, the giraffe represents not a mere collection of individual traits but a package of interrelated adaptations. It is put together according to an overall design that integrates all parts into a single pattern.”184
No one change, like an elongated neck, can appear without simultaneous changes in the heart and circulatory system. Any one change without the other and the animal will die. Thus, the theory of evolution is inadequate to explain even common and simple appearing mammals like the giraffe.
The broad array of thousands of different kinds of mammal species offer no support for the theory of evolution. If macro-evolution were true, there would be countless transitional species for each of these mammal species. For evolutionist’s claim that only the transitional fossils (1) on every continent, (2) on every island and (3) in every time period did not fossilize simply defies credibility. The vanishing transitional fossil theory seems more fitting for a magic show than a textbook.
6. Primates and Hominids: The Missing Evidence to Support Evolution.
Primates and hominids are not mentioned in the Bible. But no account of evolution would be complete without them. They allegedly prove that humans are the product of evolution.
While scientists like to assert that they make no claim about religion, many cite this alleged transitional evidence from primates to hominids to humans to prove that God did not create mankind. For example, the late Stephen J. Gould asks the rhetorical question: “Would God-for some inscrutable reason, or merely to test our faith have created five species, one after the other (Australopithecus Afarensis, Austraiopithecus Africanus, Homo Habilis, Homo Erectus, and Homo Sapien) to mimic a continuous trend of evolutionary change?”185
Another famous evolutionist, the late George Gaylord Simpson, likewise once cited this alleged transitional evidence between primates and humans to assert that: “Man is the result of a purposeless and natural process that did not have him in mind.”186
The National Academy of Sciences relies upon this alleged progression of fossil evidence to argue that the proof of evolution between primate-like creatures and humans is a closed case: “Today, however, there is no significant scientific doubt about the close evolutionary relationships among all primates or between apes and humans. The missing links’ that troubled Darwin and his followers are no longer missing.”187
These conclusions are often presented with pictures like this one below:
Source:188
But these emphatic conclusions undermine any claim that these scientists are impartial to the evidence. More importantly, modern discoveries have in fact undermined each of these claims.
The flaws in these conclusions are sevenfold: (1) primates, the first alleged link in this chain, cannot be traced to any alleged transitional species before them; (2) neither the fossil record nor DNA evidence provides proof of human evolution; (3) multiple studies have verified that Australopithecus did not walk on two feet, which was the primary reason for listing them as a transitional species leading to humans; (4) modern paleontologists have also ruled out Homo Habilis as an allegedly evolutionary transitional species leading to humans; (5) modern paleontologists have further ruled out Homo Erectus as an alleged evolutionary transitional species leading to humans (it is now called a side branch); (6) many modern paleontologists further concede that there is a lack of evidence to establish that Homo Heidelbergensis is an alleged evolutionary transitional species leading to humans; and (7) modern paleontologists have also ruled out Homo Neanderthis as an allegedly transitional species leading to humans (it is now also called a side branch).
New research in fact shows that the bi-pedal hominids that used primitive stone hunting tools (Homo Erectus/ possibly Homo Heidelbergensis and Homo Neanderthis) all served a purpose in preparing the animal kingdom for the onslaught of humans. In those continents where these hunting hominids once existed, large mammals suffered a lower extinction rate when humans arrived. Thus, God specially created these hominids to prepare the Earth for mankind.
(1) Primates: The Lack of Any Evolutionary Tree.
The oldest known primate appeared 65 million years ago.189 But if these primates formed the tree from which humans later appeared, the tree has no roots. After more than 150 years of searching, the animals which allegedly evolved into primates remain missing. Even the candidates for the predecessors to the primates have been ruled out one by one.
Evolutionists once theorized that primates evolved from some primitive insectivorous animal, like the tree shrew. Dr. Gish traced this theory back to the early writings of the 1920’s of Wilfred LeGros Clark in his studies of the Asian tree shrew Tupaia. But LeGros Clark’s early studies linking alleged behavioral similarities between tree shrews and primates have now been rejected by most evolutionists.
As early as 1966, studies by C.B.G. Campbell rejected any alleged link between the Tupaia and primates to be “unlikely.”190 More recent studies by R.D. Martin are considered by many to have disproven any link between tree shrews and primates. Unlike primates, which typically exhibit continuous maternal care to their young, the female tree shrew only visits the nest approximately 10 minutes for each 48 hour period. During this time, the female nurses the young. Moreover, the fat content of the milk of the tree shrew (approximately 25%), in no way resembles the fat content of the milk of a typical primate (normally between 1 and 3 percent). These differences and others caused Martin to conclude that “the tree shrew is not on the roster of human ancestors.”191
Without any evidence linking primates to any earlier transitional species, even ardent evolutionists concede that primates appear “out of nowhere” within the geological records. Robin McKie, an expert on primates, provides the following summary of the lack evidence: “Apes flourished and spread from Africa into Asia, between 20 to 30 million years ago, and then began a slow decline in the face of changes in the climate, which became cooler and drier. Only the lineages that led to gibbons and orangutans survived in Asia. In Africa, they evolved into humans, chimps, and gorillas. However, there is no fossil record for the last two on this list, making it very hard to guess the nature of our common ancestor. The possible candidates that have been considered include creatures called Sivapithecus, Ouranopithecus, Rudapithecus, and Dryopithecus, which are dated at about 10-12 million years old, as well as an ape known as Proconsul, which lived about 16-20 million years ago. However, none of these early apes displays all of the characteristics that we would expect in a common ancestor of humans, chimps, and gorillas. For example, those skeletons that might qualify them as the ancestor don't have the right kind of teeth, and those having the right kind of teeth and jaws have the wrong kind of skeleton - and so the mystery remains.”192
Several of the world’s other leading experts in the field of primates, have offered similar concessions: “In spite of recent finds, the time and place and origin of primates remains shrouded in mystery.193. . . the transition from insectivore to primate is not documented by fossils. The basis or knowledge about the transition is by mere inference from living forms.194 The specific origin of primates among more primitive eutherians has not been established.195 At some point, one group derived from the lower primates and gave rise to the anthropoids. But no one can say with certainty what this ancestor looked like, because there’s a large gap in the fossil record between primitive and advanced forms.”196
Dr. Gish further observed that the alleged evolutionary pathway between new world monkeys with their broader noses followed an allegedly separate pathway from old world monkeys with their narrower noses. Although each is speculated to have followed a different evolutionary pathway, in neither case can paleontologists identify any preceding species on these parallel evolutionary paths. Paleontologist A.S. Romer concedes “little is known, unfortunately, of the fossil history of the South American Monkeys.”197
Likewise, paleontologist A.J. Kelso also concedes: “The details of the evolutionary background of the New World monkeys, the platyrrhinae, would doubtless be informative and interesting, but unfortunately, we know very little about them.”198
Likewise, R.D. Martin, concedes: “The known fossil record of the New World monkeys for South and Central America is, frankly, rather disappointing in many respects . . . The origins of the New World monkeys therefore remain a mystery as the only substantial fossils are known in the late Oligocene and the Miocene, and by that stage they exhibit numerous characteristics clearly allying them with the modern forms. Indeed, without exception, the substantial fossils that have been described resemble true New World monkeys. . . .” 199
Dr. Gish quotes the same paleontologist as conceding that the same geological record also fails to provide evidence of evolution for Old World monkeys: “Clearly, the fossil documentation of the emergence of the Old World monkeys could provide key insights into the general evolutionary picture of the primate but, in fact, this record simply does not exist.200 Although early fossil evidence of New World monkeys may be described as disappointing, it is rich in comparison to that for Old World monkeys . . . Overall, the fossil record can tell us very little about the origin of New World monkeys.”201
Summarizing these quotes and others, Dr. Gish concluded that the fossil record fails to support the theory of evolution from early mammals to primates. Without any evolutionary evidence of a transition from early mammals to primates, he further concluded that the alleged evolutionary tree from mammals to primates to humans is also without evidence: “There is no evidence either in the present world or in the world of the past to link primates to any other creature. Right at the very start, then, an evolutionary origin of man is invalidated by actual empirical scientific evidence. Primates, as a group, stand completely isolated from all other creatures.”202
Thus, there is no fossil evidence linking primates to anything before them. There is also no evidence establishing that primates evolved into the hominids or humans.
(2) Paleontologists’ Failure to Find Either Fossil or DNA Evidence of Mankind’s Evolution.
After more than 150 years of searching, paleontologists have been forced to concede that the fossil record linking humans to primates is also missing. For example, paleontologist Lyell Watson, concedes that the fossil record does not support an evolutionary pathway to either apes or humans: “Modern apes, for instance, seem to have sprung up out of nowhere. They have no yesterday, no fossil record. And the true origin of modern humans of upright, naked, tool-making, big-brain beings is, if we are to be honest with ourselves, an equally mysterious matter.”203
Dr. Gish quotes R. B. Martin as offering a similar concession as to the dismal state of the alleged fossil evidence: “It should be noted at the outset that the substantial fossil remains are known for all of the species listed below (a quite unusual situation with respect to the primate fossil record generally), but that there is virtually no fossil evidence relating to human evolution, other than a few fragments of dubious affinities, before about 3.8 million years ago. The preceding period of human evolution therefore remains a complete mystery and an unfortunate major gap exists whatever view one takes of a time of divergence of hominid and great apes.”204
Paleontologist J. S. Jones and S. Rouhani offer a similar concession: “The human fossil record is no exception to the general rule that the main lesson from paleontology is that evolution always takes place somewhere else.”205
Dr. Fazale Rana, a chemist and an author on the subject, observes: “Instead of an evolutionary tree, hominid discoveries form a bush or a lawn.”206
Even the most committed advocates of human evolution concede that hominid paleontology is at best speculative evidence. As best explained by Robin McKie in the book Dawn of Man: The Story of Human Evolution (Dorling Kindersley Publishing, Inc. New York 2000) p. 26, both sides of this part of the evolutionary debate are forced to draw large conclusions from only limited evidence: “More than any other scientific profession, paleontologists have a poor reputation for squabbling . . . These disputes have occasionally become rather heated and it would be easy to conclude from them that the profession is made up of some unusually hot-headed, inflexible individuals . . . In addition, there is a distinct paucity of evidence from which to make arguments. Our understanding of our evolution is based on the discovery of the remains of only a couple of thousand cavemen or ape-men, most of whom have left a mere scrap or two of bone to posterity. In other words, there is only a limited amount of data - but a correspondingly large opportunity for speculating.”
Without proof from the fossil record, many advocates of evolution attempt to use DNA to prove human evolution. But this alleged proof also falls short. In the book “Why Evolution is True”, author Jerry Coyne concedes: “That oft-quoted 1.5 percent difference between ourselves and chimps, then is really larger than it looks … More than 6 percent of genes found in humans simply aren’t found in any form in chimpanzees. There are over fourteen hundred novel genes expressed in humans but not in chimps. … Despite our general resemblance to our primate cousins, then, evolving a human from an apelike ancestor probably required substantial genetic change.”207
Because the evidence is unclear, advocates of evolution also cannot agree on the alleged route from a single common ancestor to humans. One group believes that the alleged evolutionary branch from this common ancestor broke with our most related genetic cousins, the chimpanzee (pan and the gorilla). This group bases their claims on the similarity of DNA between man and the African apes of gorilla and chimpanzee. This group generally believes that the one percent base mismatch between the DNA of humans and chimpanzees and gorillas argues strongly that these African primates are our closest genetic cousins. These scientists argue that the alleged mismatch between the DNA between humans and orangutans is estimated to be 2.4 percent.208
The second group believes that our most related cousin is the orangutans (Pongo). Paleontologist Jeffrey Schwartz, an fierce advocate of this view, argues that the DNA evidence is overrated: “Of the 26-traits that man shares with the living Hominid, he shares all 26 with the orangutans, only 9 with the chimpanzee and gorilla, and but five with the gibbon (Hylobates). Some of the traits man shares with the orangutans but not with the African apes, according to Schwartz, include the longest hair, most widely separated mammary glands, the longest gestation period (equal to man in orangutans), thick molar enamel, low-cusped cheek teeth, copulation not confined to a specific part of the menstrual cycle, and highest oestriol levels during the menstrual cycle.”209
Thus, neither the fossil record nor DNA evidence provides proof of human evolution.
(3) Australopithecus – The Lack of Evidence Linking These Primates to Mankind.
Australopithecus is the next extinct primate that allegedly provides undisputed proof of human evolution. But the fossil record establishing that primates evolved into these creatures is also missing. Paleontologist Bernard Wood admits that the appearance of its alleged predecessor “S. tchadenis” seven million years ago is so abrupt that it is comparable to the Cambrian explosion: “The fauna of the Burgess Shale in Canada, which samples a bewildering array of invertebrate groups some 500 million years ago, is a famous example of diversity at the base of an adaptive radiation. Does S. tchadenis belong to the African equivalent of the Burgess Shale?”210
Before establishing why these creatures are unrelated to humans, a few terms need to be defined. Australopithecus is a genus classification within a “super-family” that groups together primates, allegedly intermediate creatures and humans. The super-family is called Hominoidea. Paleontologists call creatures that allegedly came between the primates and humans as “hominids”.
Australopithecus was a group of alleged hominids that are believed to have existed between approximately 4.5 million years to as recently as 1 million years. Creationists simply refer to them as extinct primates.
These primates possessed small brains with an average cranial capacity of approximately 500 cubic centimeters or less. This is the size of a typical gorilla and about 1/3 that of modern man.
In 1924, paleontologist Raymond Dart found the first of these creatures, now called Austraiopithecus Africanus. As a means of notoriety, paleontologists are frequently quick to herald each new partial fossil specimen as a new species of “Australopithecus.” This has led to classification confusion even amongst paleontologists. Under one view, there are seven documented or proposed australopithecine species. These include: (1) Australopithecus afarensis; (2) A. africanus; (3) A. anamensis; (4) A. bahrelghazali; (5) A. deyiremeda (proposed); (6) A. garhi; and (7) A. sediba.211
Yet, to add to the confusion, paleontologists cannot agree if some partial fossil specimens belong within one of these seven species or as part of a separate genus. For example, one partial fossil specimen found only in South Africa is called by some “Australopithecus robustus.” Some call it a closely related species to Australopithecus Africanus. But anthropologist Robert Broom became famous for calling this specimen “robustus” under a brand new genus that he called “Paranthropus”. It lived between 2.0 and 1.2 million years ago and possessed more massive teeth and jaws than other fossils for that time period. This primate-like species also had jaws with a more boney-like ridge, similar to that found in gorillas and orangutans.212
As another example, in 1999 Justus Erus and Meave Leakey found parts of a 3.2 to 3.5-million-year-old hominid. They called it a new genus under the classification “Kenyanthropus platyops”. But others disagreed and called it Australopithecus afarensis.213
As yet another example, some paleontologists call Australopithecus ramidus” a related species to the other Australopithecus-like primates. This species is different because it had teeth more similar to chimpanzees or other apes than modern humans. Fossils from this species are 4.5 million years old, a ½-million years older than Australopithecus Africanus.214
But the wide diversity of names given to these creatures does not prove that they evolved from anything. They all lived concurrently and therefore could not have evolved from each other. There is also no fossil evidence that the genus Australopithecus or the other similar creatures evolved from primates. More importantly, the primary reason for alleging that they are mankind’s ancestor (based upon an allegedly human-like bipedal gait) has been disproved in multiple studies.
Modern computer studies have repeatedly ruled out the possibility that Australopithecus had a human-like bipedal gait. But because many textbooks continue to publish that false claim that these extinct primates walked like humans, the details of these studies are discussed here in detail.
One famous study was done by the British Anatomist, Solly Lord Zuckerman and Dr. Chades Oxnard, former director of graduate studies and professor of entomology at the University of Southern California Medical School. For over 15 years, Lord Zuckerman led a comprehensive research team studying the anatomical features of the Australopithecus fossils as well as fossils from apes, humans, and other forms of primates. From these studies, Oxnard concluded that Australopithecus did not walk in an upright manner like humans: “Multivarite studies of several anatomical regions, shoulders, pelvis, ankle, foot, elbow, and hand are not available for the Australopithecus. These suggest that the common view that these fossils are similar to modern man or that those occasions when they depart from the similarity to man they resemble the African great apes may be incorrect. Most of the fossil fragments are in fact uniquely different from both man and man's nearest living genetic relatives, the chimpanzee and the gorilla.”215
Oxnard later conclusively rejected the alleged evolutionary pathway from Australopithecus to humans: “The Australopithecine known over the last few decades . . . are now irrevocably removed from a place in the evolution of human bipedalism possibly from a place in a group any closer to humans than the African apes and certainly from a place in the direct human lineage.”216
Based upon the same evidence, Lord Zuckerman also rejected any claim that Australopithecus should be classified as an alleged transitional species from some early primate to man: “But I myself remain totally unpersuaded. Almost always when I tried to check the anatomical claims on which the status of Australopithecus is based, I have ended in failure.”217 He concluded that Australopithecus was an ape in no way related to the origin of man.
Another comprehensive study by two other anthropologists, Stern and Susman, also established that these creatures were primarily arboreal and did not walk in a bipedal manner similar to humans: “A summary of the morphologic and functional affinities of the hadar (the geographic location where these fossils were found) hand fossils, heads inexorably to an image of a sensory adapted hand, surprisingly similar to hands found in the small end of the pygmy chimpanzee-common chimpanzee range. . . In summary, the foot and ankle remains reveal to us an animal that was engaged in climbing as well as bipedality . . . There is no evidence that these extinct primates had long curved heavily muscled hands and feet for any purpose other than to meet the demands of full or part-time arboreal life.”218
Two famous paleontologists, Ian Tattersall and Jeffrey Schwartz, have recently offered even more definitive proof that these primates did not walk like humans. Unlike most paleontologists who only study casts of the original remains, Tattersall and Schwartz inspected nearly every known original hominid fossil remain throughout the world in preparation for their study. The two based their comprehensive conclusions on several important factors. First, the two agreed with the research of paleontologist Peter Schmid, who reconstructed the pelvis to be broader from side to side and more turned out above the hip joints. Schmid also reconstructed the rib cage to be more conical in structure, narrow at the top and fanning out considerably at the bottom. This reconstruction is now accepted by other noted paleontologists, even if all of the conclusions that follow are not: “In apes and australopithecines the rib cage is shaped like a pyramid that gets larger as you move down the body ...”219
As a result of this reconstruction, Tattersall and Schwartz agreed with Schmid’s conclusions that these creatures were considered less likely to have walked upright. Second, the two noted that footprints found at Laetoli, Tanzania, which were previously thought to be evidence of animals with a bipedal gait, suggest otherwise. Paleontologist Ron Clarke found that chimpanzees leave similar depressions in damp soil when they walk on their hind feet. Summarizing these factors and others, Tattersall and Schwartz conclude that these creatures were not bipedal as previously thought: “[T]he arms of the afarensis were still quite long. Less debatable is the fact that the legs of afarensis were quite short. As for the fingers and toes, they were quite long, and the shafts of these bones were curved (as in orangutans and gibbons) in contrast to humans and even to the terrestrial gorillas (in which these bones are relatively short and their shafts rather straight). In addition, the groove along the backside of the lower end of the outer lower leg bone, the fibula, through which the tendon of a muscle inserting on the sole of the foot passes, is not only deeper than in humans, but also deeper than in chimpanzees. And this, in turn suggests that the larger muscle of afarensis would have been better able to stabilize its foot, as when being pressed against a vertical support, than in any human or chimpanzee. Put these afarensis features together and, according to Randy Susman and colleagues at SUNY-Stony Brook, you have an australopith that was probably more committed to life in the trees than on the ground. In fact, the Stony Brook group concluded, many features that had come to be associated with bipedalism may very well have been useful to afarensis (and, by extension, other australopiths) for [moving] up and down trees. Only when these hominids had to travel between arboreal habitats would they find themselves walking bipedally, “forced” to do so by their own anatomies.”220
Two studies in the year 2000 backed up these findings. These studies also concluded that australopiths did not walk on both feet. Instead, they knuckle walked like the great apes.221
Tattersall, Schwartz, Clarke, Schmid, Stern, Susman, Zuckerman, and Oxnard each believe in the theory of evolution. But in systematically dismantling the claims that the Australopithecus walked like humans, these paleontologists dismantle the primary reason for placing these creatures on the alleged evolutionary pathway related to humans. In reaching their findings, they undermine the very claims of those who refer to these creatures as proof of the theory of evolution.
Most paleontologists further agree that Australopithecus did not use tools of any kind. Only one study claims that a primate called Australopithecus garhi “may” have used crude instruments to remove flesh from an animal’s remains.222 That is it.
These animals possessed small brains with an average cranial capacity of a typical gorilla, approximately 500 cubic centimeters or less. These creatures were nothing more than extinct primates. In fact, the Greek word “Australo” and the Latin word “-pithecus” literally means “southern ape.” They were exactly what their name suggests.
(4) Homo Habilis – The Lack of Evidence Linking These Hominids to Mankind.
Hominids in the genus “Homo” formed the next alleged evolutionary link from Australopithecus to mankind. But there is no agreed upon definition for this genus. After looking at every hominid fossil, Tattersall and Schwartz conceded: “There is, thus, still no agreed-upon morphological definition for the genus Homo.” More importantly, the two concede that there is no fossil evidence that the poorly defined genus Australopithecus evolved into the poorly defined genus Homo: “the general pattern is clear. The million years or so after the 2.5-[million year] mark there is no simple linear transition from one species of gracile Australopithecus to a successor species of Homo.”223
In addition to having no evidence that genus Australopithecus evolved into the genus Homo, paleontologists have no idea which species of the genus Homo allegedly first evolved from Australopithecus. Until recently, a species called “Homo Habilis” was touted as the first link in the chain. As quoted at the beginning of this section, the late Stephen J. Gould cited this as one of “five species” immediately after “Austraiopithecus Africanus” that formed “a continuous trend of evolutionary change.”224
Paleontologists even created compelling pictures showing an allegedly smooth transition from the Australopithecus-related species to Homo Habilis. Homo Habilis was slightly larger in size. It also had a cranial capacity of an estimated range between 650 and 775 cubic centimeters. Fossil records of this species are carbon-dated to a range between 1.7 and 1.9 million years ago. However, no complete skeletons or skulls exist for this group. Moreover, the slightly larger size proves nothing. There are apes of multiple different sizes today.
Furthermore, paleontologists are now questioning whether this once allegedly critical link in the chain to human evolution was even a separate species. After examining every known hominid fossil, Tattersall and Schwartz concluded that Homo Habilis was not an alleged evolutionary descendant of Australopithecus. The two even called Homo Habilis a “wastebasket species.”225 They further concluded: “And it has to be admitted that the new hominid itself consists of more than a couple of handfuls of fragments that could be interpreted in several different ways.”226
In addition to the lack of any evidence of evolution between the genus Australopithecus and the genus Homo, paleontologists have also verified that the two genus groups lived at the same time. Richard Leakey and Alan Walker, both noted paleontologists, have verified that: “There is evidence from East Africa for a late-surviving small Australopithecus individuals that were contemporaneous first with Homo Habilis, and then with Homo Erectus.”227
Likewise, Dr. Gish quotes the magazine Nature as offering the additional concession regarding the contemporaneous existence of Australopithecus and Homo Habilis: “Such sympathy for Homo with one, and quite possibly two forms of Australopithecine could mean that there is little information about the direct and intermediate ancestors of Homo. These known Australopithecine, contemporaneous with Homo, obviously cannot fill the ancestry role.”228
For these reasons, many paleontologists, like Robert Boyd and Joan Silk, argue that Homo Habilis along with another group called Homo Rudolfensis that coexisted with them were really members of Australopithecus.229 Thus, these were simply extinct primates that form no part of an alleged evolutionary tree. The chain that allegedly tested mankind’s faith is disappearing with each new scientific discovery.
After boasting of the evolutionary tree from primates to humans, even the late Stephen J. Gould was forced to concede before his death that Homo Habilis cannot form an evolutionary link between Australopithecus and mankind: “What has become of our lineage if there are three coexisting lineages of hominid (Australopithecus Africanus, the robustus Australopithecine, and Homo habilis) none clearly derived from another? Moreover, none of these display any evolutionary trends during their tenure on Earth. None became brainier or more erect as they approached the present day.”230
Paleontologist McKie likewise quotes the famous paleontologist named Meave Leakey to argue that Homo Habilis has no clear role in our alleged evolutionary pathway: “‘The trouble is that we lack good evidence. We desperately need a good skull and skeleton of a single member of the species to resolve this problem. Certainly this stage of human evolution is one of the most difficult to understand.’ Habilis, in short, is a problem.”231
There is some limited evidence that Homo Habilis used rock flakes as a tool.232 But many primates today use sticks as tools when needed to obtain food. This does not establish that they were evolving to become more like humans in either their intellect or consciousness.
Thus, neither Australopithecus nor Homo Habilis offer any support for the allegedly definitive proof of evolution between primates and humans. Evolutionists are fond of telling those who believe in the Bible to wait for science to prove the claims of evolution to be true. But the exact opposite is happening. The more we learn about these extinct primates, the more we learn that the theory of human evolution is unsupported by anything in the fossil record.
(5) Homo Erectus – The Lack of Evidence Linking These Hominids to Mankind.
The late Stephen J. Gould offered “Homo Erectus” as the last species that formed a “continuous trend of evolutionary change” leading to Homo Sapien.233 Until recently, paleontologists claimed that they had evidence of continuous change between Homo Habilis to Homo Erectus to Homo Sapien.
Paleontologists were quick to point out that Homo Erectus was slightly larger than Homo Habilis, just what their evolutionary model predicted. They had a cranial capacity between 800 and 850 cubic centimeters. This species is alleged to have existed from approximately 1.8 to 1.5 million years ago. They also used primitive rock flakes as tools. They further hunted with sharp sticks. They received the name Homo Erectus because it is claimed that they had the ability to walk erect. Thus, paleontologists once called them our ancestors.
But the once unquestionable evidence showing that Homo Erectus evolved into humans has crumbled with closer scrutiny. Homo Erectus is now considered by most paleontologists to be an evolutionary side branch that arose in Asia. As a result of their comprehensive examination and study of every known hominid fossil, Tattersall and Schwartz were also forced to reject Homo Erectus as an alleged common ancestor to humans: “[W]e clearly have to abandon the view that H. Erectus represented the main line of human evolution in the run-up to the appearance of H. Sapiens. In other words, we have to step aside from the linear paradigm that has governed recent investigations in paleoanthropology. Instead, we need to accept that H. Erectus was actually an indigenous and terminal eastern Asian development: simply - just like the Neanderthals thousands of miles away - another twig on the hominid evolutionary bush that was eventually pruned.”234
Tattersall and Schwartz instead suggest that Homo Ergaster was the alleged next common descendant after Australopithecus. But other paleontologists like Robin McKie have rejected this conclusion as being unsupported.235
Only one near complete hominid skeleton exists, called “the Nariokotome boy.” (see below) In nearly every other case, paleontologists are forced to make assumptions about how the bones were to be reconstructed and what the hominids looked like.
Source:236
The contributors to Wikipedia (source of picture above) call this Homo Ergaster.237 But paleontologists McKie and Alan Walker (who discovered the skeleton), call it Homo Erectus in the quote below. Because the genus Homo has no clear definition, paleontologists cannot even agree what to call it.
Whatever it should be called, this nearly complete fossil allows for humans to test whether these hominids (Homo Erectus / Homo Ergaster) became more human-like in their intelligence. In fact, the Nariokotome boy skeleton establishes that these creatures were not becoming like humans in either intellect or consciousness. After an extensive study, paleontologists McKie and Walker, conceded that these were creatures without either the ability to speak or a human-like consciousness: “[I]t was the study of the Nariokotome boy’s head and upper parts that were to provide the most notable and startling results. Initial measurements showed that his brain must have had a volume of only 880 cc - about double the volume of a typical ape and about two-thirds of the modern human average of 1350 cc. It may sound like a fairly respectable cranial capacity, but Walker disagrees. Compared with modern humans, the Nariokotome boy ‘had the height of a 15-year-old with the brain of a one-year-old.’ Not quite so impressive, in other words. . . But the real shock came when scientists studied the Nariokotome boy's spine. Ann MacLarnon, an anthropologist at the Roehampton Institute in London, found that the vertebral canal - a hollow passage in the spine that carries nerves from the brain to the rest of the body - was narrower near the chest than it is in modern humans. Together with other evidence, this suggested that Homo Erectus did not have fine control over the muscles involved in breathing - muscles that we employ, quite unconsciously, in speech. Language, as we understand it, has probably not yet evolved. . . He had not mastered language, and that meant he was far from possessing what we understand as ‘human spirit’. As Walker puts it, ‘in his eyes not the expectant reserve of a stranger but that deadly unknowing I have seen in a lion’s blank eyes. He may have been our ancestor, but there was no human consciousness within the human body. He was not one of us.’ Starved of intellect, the Nariokotome boy could not talk, and he could only have been able to make crude mental maps of his world.”238
Paleontologist Desmond Clark of the University of California at Berkeley likewise concludes: “If these ancient people were talking to each other, they were saying the same thing over and over again.”239
They also showed no cultural development to suggest that they were becoming like humans. They did not have clothes, art, jewelry, pottery, a written language or complex tools.
Moreover, to the extent many believe that Homo Erectus evolved, there is no evidence of where or what it evolved into: “Unlike so many other lineages that we have discussed in this book, extinction was not to be its fate. It would evolve, although exactly where and into what is the intriguing question.”240
Thus, there is no evidence that Homo erectus evolved into humans. They were simply one of many species of extinct primates or hominids.
Other Homo creatures that have been nominated to fill the void include: Homo ergaster, Homo rudolfensis, Homo steinheimensis (for which only one partial skull exist and which is considered by many to be misclassified)241, Homo Naledi (for which only 15 partial fossils exist in one cave)242, and Homo Heidelbergensis (discussed below). But the claims supporting each of these species have also fallen short. As quoted previously, paleontologists Boyd and Silk established that Homo rudolfensis were really members of Australopithecus.243 Homo ergaster is either a misnamed Homo erectus or another alleged evolutionary side branch of primate-like hominids.
(6) Homo Heidelbergensis – The Lack of Evidence Linking Them to Mankind.
Homo Heidelbergensis, also called Homo rhodesiensis, is now offered by some paleontologists as the alleged common ancestor between Homo Neanderthis and humans. They lived in Africa, Europe, and western Asia between 600 and 200 thousand years ago. They had a typical cranial volume of 1100–1400 cm³. Humans have an cranial volume of 1350 cm³.244
Paleontologists, however, have been limited to a sample size of approximately 50 partial hominid remains from Homo Heidelbergensis. Moreover, 32 partial remains of this total are found in one cave called Atapuerca Cavern in Northern Spain.245 The actual number of hominids at issue is even smaller. Thus, too little is known about this group to draw sweeping conclusions in either direction. Like other hominids, its alleged predecessor in the fossil record remains unknown.246
Robin McKie argues that the limited fossil sites contain few clues about this group: “Unfortunately, there are not enough fossils to give a clear picture of the heidelberg anatomy. For example, only a couple of molar teeth and a shinbone were found at Boxgrove [England].”247
With only 50 partial fragments, there are serious questions about whether Homo Heidelbergensis is properly classified as its own species. Even if it were a separate species, the alleged evidence linking Homo Heidelbergensis to humans is missing.
Assuming they are properly classified, they also showed no signs of human intellect or consciousness. They did not have clothes, art or sophisticated tools. They only had stone tools allegedly used by other hominids. They further did not have a spoken language. Paleontologist Steven Mithen believes that they had a “pre-linguistic system of communication.”248 Of course, all animals have a non-linguistic system of communication. This only confirms that it did not represent an evolutionary step toward mankind.
(7) Homo Neanderthis – Evidence That Mankind Did Not Evolve from Them.
Finally, Homo Neanderthis was also once called ancestral to mankind. Unlike the limited partial fossil remains for the species Homo Heidelbergensis and Homo Naledi, many fossils have been found for Homo Neanderthis. The first two fossils were found in Europe in 1829 and 1848. But people at the time did not realize what they were. After a third fossil was found in 1856 in the Neander valley, 12 km (7.5 mi) east of Düsseldorf, Germany, they obtained the name “Neanderthal.” At the time, many advocates of Darwin’s theory of evolution heralded them as the missing link and proof of mankind’s evolution.
But these claims have also been disproven. Fossil evidence showing that Homo Neanderthis lived both prior to and concurrently with humans establishes that mankind did not evolve from them. Based upon recovered fossils, they overlapped with human populations throughout all of Europe and portions of the Middle East and Asia Minor. Paleontologists now call them an evolutionary off-shoot to the presently unknown alleged predecessor to our own species. 249
Establishing that humans didn't evolve from Homo Neanderthis, however, does not solve the question regarding why they existed. It is the very fact that they existed that gives many reasons to believe in evolution. They were very different from the other hominids. They had larger brains than humans. The average cranial capacity of Neanderthals was 1600 cm3. This was much larger than the 1400 cm3 average for modern humans. Males stood 164–168 cm (65–66 in) and females 152–156 cm (60–61 in) tall.250 Some researchers even claim that they were stronger than humans.251 Some further claim that their eyesight was better than humans.252

(Neanderthal skull at La Chapelle-aux-Saints France in 1908)253
Some have also claimed that they used stone tools that were more advanced than the hominids before them. Some 300,000-year-old tools predated humans and are attributed to them.254 Other stone tools found in a cave on the south-facing coast of Gibraltar are also attributed to them.255 Some scientists have speculated that they may have even used watercraft similar to dugout canoes.256 Certain stone tools discovered on the southern Ionian Islands have even caused others to speculate that they were sailing the Mediterranean Sea as early as 110,000 years ago.257 Other 170,000-year-old quartz hand-axes, three-sided picks, and stone cleavers on Crete further caused some to speculate that they sailed to that island as well.258
Some researchers have even claimed to have found fossilized grains of allegedly cooked vegetable material in their teeth.259 Thus, some claim that they also used fire.
One paleontologist has claimed that they buried their dead with flowers. But this is based solely upon the existence of pollen near recovered Neanderthal bones.260 Pollen is easily disbursed and could easily have contaminated a dead Neanderthal body. Thus, this evidence is not compelling.
But regardless of whether they buried their dead with flowers, they were different from the hominids. The unstated feeling of many is that God had no reason to create them if He only created humans in His image. Or, if He did, He would have mentioned them in the Bible.
Mankind, however, can never know God’s mind. When Job wrestled with God’s actions, God responded: “Where were you when I laid the foundation of the earth? Tell Me, if you have understanding,” (Job 38:4). Thus, God does not provide answers to every question that we ask.
Nevertheless, creationists have developed three possible theories to explain them. Under one theory, they were the mysterious “Nephilim” in the book of Genesis. (Gen. 6:4). Commentator John Denton provides several arguments to support this view.261 He claims that the Nephilim were the “giants” who bred with mankind against God’s will. God later wiped them out in the flood.
He suggests that Cain married a female Neanderthal. The Bible states that Cain took a wife and had children in the land of Nod after he was expelled following his murder of his brother Able. (Gen. 4:16-17). At the historic Scopes trial in Tennessee in 1925, an outspokenly anti-Christian ACLU lawyer named Clarence Darrow cross-examined prosecutor William Jennings Bryan about the origin of Cain’s wife. When Bryan could not provide an answer, the press claimed that Christians were unable to defend the biblical record. Or, they were the people Cain feared at a time when God drove him out of the Garden of Eden and no other men besides Adam were yet alive.
Initial studies found that the Neanderthals did not breed with humans. But later more comprehensive studies found that they did.
In 1997 and in 2000, geneticists were able to extract Mitochondrial DNA sequences from three different Neanderthal specimens in three different regions from bones. The studies found no sign of modern human descent from the Neanderthals.262
In 2006, the Lawrence Berkeley National Laboratory found that Neanderthals and humans most likely did not interbreed.263 Edward M. Rubin, director of the U.S. Department of Energy’s Lawrence Berkeley National Laboratory and the Joint Genome Institute (JGI), declared: “While unable to definitively conclude that interbreeding between the two species of humans did not occur, analysis of the nuclear DNA from the Neanderthal suggests the low likelihood of it having occurred at any appreciable level.”264
In 2008, the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, published the full sequence of Neanderthal mitochondrial DNA (mtDNA). It also found that Neanderthals “probably did not interbreed with their human neighbors.”265
But the Neanderthal genome project published in 2010 and 2014 reached a different conclusion after completely sequencing the entire Neanderthal genome. It found that Neanderthals contributed to the DNA of most non-Africans humans and a few African populations through interbreeding between 50,000 to 60,000 years ago. The study claims that between 1.5 and 2.1 percent of the DNA of non-African humans is Neanderthal in origin.266
The Neanderthal-Nephilim interpretation, however, is currently the minority view amongst Bible scholars. This may, however, be partially attributed to the relatively recent DNA findings.
The majority of Bible commentators argue that the Neanderthals could not be the Nephilim because that same Hebrew name is used to refer to the ‘giants' that the Jews encountered centuries later in Canaan (Nu. 13:33). A similar Hebrew word with different vowel-sounds is also used to refer to dead Philistine soldiers (Ez. 32:27). Most Bible commentators further claim that Cain married one of his sisters.
Those who hold to Denton’s view respond that the term Nephilim could refer to both Neanderthal giants and later Canaanite giants. Hebrew has far fewer words than in English with many words having multiple meanings. Under this view, the Neanderthal DNA would still be present in human DNA today through one of Noah’s son’s wives on the ark.
Under an entirely different second view, the Neanderthals were allegedly humans. Many young earth creationists, like Dr. Duane Gish, hold this view. Many claim that evidence of interbreeding supports this view because the standard definition of a species is a group that can interbreed with its own members.267 But their DNA differed from human DNA. Moreover, as set forth in the next section, humans were qualitatively different from Neanderthals in every way. Examples include tools, weapons, clothes, art, jewelry, social structures, longevity, the ability to innovate, burial practices, and worship.
Hugh Ross provides a third explanation for both the Neanderthals and the other bipedal hominids that hunted before them. He argues God most likely created them to prepare the animals for the arrival of mankind. The Neanderthals, for example, were apex predators that were capable of bringing down a wide range of prey from reindeer, red deer, ibex, wild boar, aurochs, mammoth, elephants, and rhinoceros.268 Animals in places where hominids lived before the arrival of mankind had far higher survival rates than places where mankind arrived on the scene without any prior hominid species present: “In Australia, where fossil evidence suggests no hominid species resided prior to humans, only 1 out of 16 large-bodied mammal genera survived the arrival of humans. By contrast, in sub-Saharan Africa where the full panoply of hominid species fossils have been discovered, only 2 out of 44 large-bodied mammal genera went extinct upon modern humans’ arrival. Apparently, increasing exposure to the predatory skills of successive hominid species allowed birds and mammals to prepare for the shock of a superior predator.”269
In places where the humans did wipe out the large mammals, they later suffered: “In those continental regions that lacked bipedal primate species previous to humans and where most of the large-body mammal species were wiped out shortly after the arrival of humans (Australia, South America, and North America) humans there advanced little, if any, beyond a stone-age existence. Not until the arrival of both people and large-bodied mammals from Europe and Asia in the sixteenth century was this limitation overcome.”270
Thus, neither the Neanderthals nor the hominids arrived by chance. God created them to prepare the animals of the Earth for His greatest (but also most destructive creation) mankind.
7. Humans: Evidence of God’s Special Creation. Gen. 1:26-31
In the final part of Genesis chapter one, Moses revealed that God created mankind as His last creation (Gen. 1:26-31). Moses again made a revelation that has again been proved true. With the possible exception of new strands of bacteria or viruses, no new species have appeared on the planet since Moses. Again, there would be no way for Moses on his own to predict this fact.
Through the third and final usage of the Hebrew word bara, he further revealed that God created man from literally nothing. (Gen. 1:26-27). “Created.—This significant verb is thrice repeated with reference to man. It indicates, first, that man has that in him which was not a development or evolution, but something new. He is, in fact, the most perfect work of the creative energy, and differs from the animals not only in degree, but in kind, though possessing, in common with them, an organised body. And next, it indicates the rejoicing of the Deity at the completion of His purpose.” (Charles Ellicott’s Commentary on Gen. 1:26-28).271 In the next chapter, he reveals that God created man from the “dust” (Gen. 2:7). Thus, mankind did not evolve from anything else.
The Bible further reveals that God made mankind in His image (Genesis 1:27). “Man was to be a creature different from all that had been hitherto made. Flesh and spirit, heaven and earth, must be put together in him. God said, Let us make man. Man, when he was made, was to glorify the Father, Son, and Holy Ghost. Into that great name we are baptized, for to that great name we owe our being. It is the soul of man that especially bears God's image.” (Matthew Henry on Gen. 1:27).272 Although mankind cannot fully know all that that entails, it includes at least seven things that distinguish it from anything else that God created. These include: (1) mankind can use complex language with abstract thought and grammar; (2) mankind has a unique ability for self-expression, creativity, and innovation; (3) mankind can form complex interdependent social networks; (4) mankind has the ability to worship and concern itself with spiritual matters; (5) mankind can concern itself with matters of morality and justice; (6) mankind can concern itself with matters of governance and stewardship; and (7) mankind has a unique ability to show love and compassion toward others.
(1) Mankind’s Unique Complex Language Abilities.
All animals communicate with others within their same species. But one of the most important differences between mankind and the animal kingdom is its unique ability to communicate abstract thoughts through complex forms of spoken and written language. This also allowed it to dominate, organize, innovate, and progress as a special species created in God’s image.
Consistent with mankind’s unique status, decades of studies on the language ability of modern primates has failed to establish any ability in our alleged genetic cousins to think in abstract terms. This also allows humans to express themselves in over 5,000 different languages.
Mankind’s qualitative language advantage over the hominids. As stated previously, the near complete skeleton of the Nariokotome boy allowed for scientists to test the language abilities of Homo Erectus / Homo Ergaster. Paleontologists McKie and Walker verified that these were creatures without either the ability to speak or a human-like consciousness: “Language, as we understand it, had probably not yet evolved.”273 Paleontologist Desmond Clark of the University of California at Berkeley reached the same conclusion.274 Paleontologist Steven Mithen further found that Homo Heidelbergensis and the Neanderthals at best had a “pre-linguistic system of communication.”275 Thus, there is no evidence that humans developed language through evolution from these hominids.
Mankind’s qualitative language differences over primates. Two noted linguists, John W. Oller, Jr. and John L. Omdahl, confirmed the uniqueness of human abstract thought in a comparison study of twenty years of research related to the limits of language ability in our alleged primate cousins.276 Oller and Omdahl conclude, “human beings are uniquely equipped to acquire languages and our capacity to do so involves a remarkably abstract system of expression that is totally unparalleled in any other species.”277
The inability of primates to ask questions. In one study which began in 1966, researchers sought to teach chimpanzees the American Sign Language (ASL) for the deaf. In the end, the chimpanzees acquired approximately 130 words which they used with an average accuracy rate of 80%.278 By 1978, a Stanford psychologist, Francine Patterson, had taught a famous gorilla named Koko to use approximately 375 sign language symbols over a six and one-half year training period.279 By 1981, Koko was alleged to have learned approximately 600 sign language symbols. Many of her symbols she quickly forgot. Nevertheless, she was widely publicized for having engaged in such human-like dialogue as cursing, arguing with her trainers, pretending to smoke, lying about poking holes in a screen mesh on her trailer house, holding pretend tea parties, and reacting to childhood stories with all kinds of humor.280 These studies reaffirmed what many researchers already knew, primates, like dolphins and whales, are highly intelligent. Nevertheless, these studies also verified the boundaries of their ability for intellectual abstract thought. None of the primates were ever able to ask questions of their own. The failure to ask questions showed limits in their cognitive reasoning abilities.
The primates inability to learn grammar. The primates were also able to learn the rules of grammar. “The absence of questions is paralleled by a complete absence of the sort of dependence on syntactic structure (rule-governed arrangement) that is universal to all human language systems and that is relied upon by all normal children in the process of acquiring their first language.”281
The primates inability to engage in “pragmatic recursion”. The primates further lacked the ability to engage in the principle of “pragmatic recursion.” This is the ability to talk about talk, such as “let’s have a story.” Although little children are capable of such comments, primates are not. Oller and Omdahl note, “Koko never argues about the meaning of signs or their appropriateness, yet every child is prone to do so.”282 Finally, primates are limited in their ability to learn language-related symbols. Oller and Omdahl note, “unlike the case of the trained ape, who achieves only about an 80% level of accuracy using signs, the child, even at playing, rarely applies a sign that is not articulately appropriate to the situation to which it is applied.”
The Bible reveals that language comes from God. The first humans all spoke a single-God given language. But God demonstrated His control over and sovereignty over human language after they built the tower of Babel. God then created mankind’s 5,000 different languages and scattered them across the Earth (Gen. 11:1-9).
Mankind’s unique language abilities and ability for abstract thought in turn allowed them to dominate over the Neanderthals and the world. This also made other innovations possible.
(2) Mankind’s Unique Ability for Abstract Expression, Creativity, and Innovation.
As another consequence of being made in God’s image, humans have the unique ability to express themselves through abstract expression, creativity, and innovation. This is evidenced by mankind’s explosion of art, technology, and innovation in the fossil record.
The human explosion in tool technology. Between 40,000 and 50,000 years ago, never before seen human innovations with tools ``exploded” into the fossil record. According to Dr. Fazale Rana, “In addition to employing stone, the first humans used ivory, bone, and wood. They transported the raw materials used for tool production at significant distances. Stone tools became smaller in size and were with more precision than those recovered from the Middle Stone Age and other comparable ancient sites. Often these later stone implements were intentionally dulled at one end and attached to wooden handles and shafts. Other tools were cut from bone and ivory, then carefully carved and polished. The first humans made projectile points, awls, punches, fish hooks, harpoons, and needles with eyes. They lived in solidly built dwellings, and made fireplaces and hearths bordered with stones for heat retention. Compared to the earlier hominids, the first humans behaved in sophisticated ways that reflected superior cognitive abilities and technical inventiveness.”283
The human explosion in hunting technology. At this time, humans also developed new kinds of nets, boats, and advanced weapons like spears and axes to capture fish and kill large animals.284
The human explosion in jewelry technology. Around this same time period, humans began to use jewelry in ways that had never been seen previously. Also according to Dr. Rana, “Unlike the hominids that predate them in the fossil record, the first humans invested time and energy in making jewelry and other items of personal adornment. They used teeth, bone, antlers, amber, ivory, marine shells, polished stones, and ostrich eggshells to make beads, rings, and pendants.”285
The human explosion in cave art. Art dating back to 25,000 B.C. or earlier, cave and rock paintings have been found in southern Africa, Europe, Siberia, India, and Australia. The first artists used stone (flint, obsidian), wood, or bone tools. Through their ingenuity, they discovered that they could use iron oxide to paint in red, and manganese oxide to paint in black.286 According to Dr. Rana, there is no record of any type of art amongst the hominids in the fossil record.287
The human explosion in musical instruments. During this time period, musical instruments also exploded wherever humans were present. Also according to Rana, “Artistic and musical expression was not part of the earlier hominids’ life. This behavior is unique to humans and coincides exclusively with human remains. Perhaps one of the most important advances in prehistoric archeology in recent years is the recognition that artistic (including musical) expression didn’t gradually emerge but rather exploded onto the scene simultaneously with humanity’s appearance about 40,000 years ago. Archeologists and anthropologists refer to this surge of human culture as ‘the big bang of art.’” 288
The human explosion in clothing technology. Around the same time period, clothing and ivory sewing needles appeared. Also according to Dr. Rana, “Clothing use also appears to be a practice associated exclusively with humans.”289
The Bible’s revelation that creativity comes from God. The Bible reveals that human creativity is a gift from God: “I have filled him with the Spirit of God in wisdom, in understanding, in knowledge, and in all kinds of craftsmanship,” (Ex. 31:3). “Now there are varieties of gifts, but the same Spirit.” (1 Cor. 12:1). “God also testifies with them, both by signs and wonders and by various miracles and by gifts of the Holy Spirit according to His own will.” (Heb. 2:4).
Some speculate that mankind appeared as far back as 200,000 years ago.290 But Dr. Rana notes that: “Between about 40,000 and 80,000 years ago, humans are largely nonexistent in the fossil record . . .” 291 Hugh Ross suggests that “Noah would have been alive roughly 40,000 years ago and Adam and Eve anywhere from 60,000 to 100,000 years ago.”292
(3) Mankind’s Unique Ability to Form Complex Social Networks.
As another consequence of being made in God’s image, humans have the unique ability to form complex social networks. Mankind’s ability to live together in large societies gave it advantages that allowed for all kinds of other advancements.
Large social networks. Animals can and do form social networks. But these networks are usually far smaller in size and limited to a pride, a colony, or a family unit. By contrast, humans have the unique ability to form social networks of unlimited size that are not based upon blood relations. They also have the unique ability to form multiple different networks at the same time. This also allowed for many types of changes and innovations. By way of comparison, researchers have concluded that Neanderthals lived in much smaller and more sparsely distributed groups than their human contemporaries. It is believed that human communities outnumber Neanderthal communities by as much as 9 to 1. Under current thinking, this allowed humans to drive out and / or kill off the Neanderthals.293
Increased longevity. Mankind’s unique ability to form large and complex social networks also helped to increase mankind’s longevity. According to Dr. Rana, “Before the first humans appeared, few hominids lived much beyond reproductive maturity. With the advent of human beings, however, longevity in the population dramatically increased.” 294 Humans began to care for the sick, the young, and the elderly in communal settings. They were also better able to protect themselves. These factors allowed human populations to explode and increase in longevity.
Specialization. As mankind formed large complex social networks and collectives, members could specialize. In the beginning, some gathered food, some cooked, some guarded the people, and others ruled. The ability to specialize in turn allowed for the pace of technological innovation to quicken. It also allowed for leisure activities like music and art.
Agriculture and villages. Beginning approximately 12,000 years ago, the Neolithic revolution took place. This included the domestication of plants through agriculture and animals through farms. This allowed society to be transformed from small, mobile groups of hunter-gatherers into sedentary societies in villages and towns. This form of stable and increased food supply and calories in turn created a further population explosion, specialization, innovation, increased longevity, and new forms of governance
The Bible reveals that the human desire for companionship comes from God. God created the uniquely human desire for companionship and friendship shortly after He made Adam: “Then the LORD God said, ‘It is not good for the man to be alone; I will make him a helper suitable for him.’” (Gen. 2:18). “Bear one another’s burdens, and thereby fulfill the law of Christ.” (Gal. 6:2). Just as God never leaves His people, His people are not to abandon each other: “The LORD is the one who goes ahead of you; He will be with you. He will not fail you or forsake you. Do not fear or be dismayed.” (Dt. 31:8; Heb. 13:5).
(4) Mankind’s Unique Ability to Concern Itself with Spiritual Matters.
As another consequence of being made in God’s image, humans have the unique ability to concern themselves with spiritual matters. We alone have complex systems of worship. We alone concern ourselves with matters of life after death and God’s existence. Mankind’s concern with spiritual matters also separates it from the hominids that preceded it.
Burial practices. Neanderthals buried their dead. Yet, according to Dr. Rana, “these burial practices appear non-ritualistic and very simple. The Neanderthals dug shallow graves that contained few if any artifacts. Human burial practices contrast sharply. Often multiple burial plots are found together. Occasionally they appear to comprise a graveyard or cemetery. Large rocks covered some graves. Such stones may reflect ritual behavior and also a desire to protect and preserve the human body. Early burials involved a profusion of grave goods (special items and body ornaments), providing strong evidence for ritualistic beliefs. These articles suggest that humanity had a sense of an afterlife.” 295
Universal worship across early societies. If spirituality were simply the product of evolution, one would expect to see some groups of indigenous societies without systems of worship and beliefs in deities. But without any documented exceptions, indigenous societies throughout the world have all independently formed systems of worship. This remained true for tens for centuries. Before the French Revolution (1787-1789) and the later writings of Karl Marx (1818-1883), atheism had no serious or organized following. It was so rare in Biblical times that there is only one verse that addresses atheism: “The fool has said in his heart, ‘There is no God.’ They are corrupt, they have committed abominable deeds; there is no one who does good.” (Ps. 14:1).
God created mankind to worship Him. Mankind’s universal desire to worship is part of our being made in God’s image. He created mankind to fellowship with Him through worship: “Everyone who is called by My name, and whom I have created for My glory, whom I have formed, even whom I have made.” (Is. 43:7). “Know that the LORD Himself is God; it is He who has made us, and not we ourselves; we are His people and the sheep of His pasture.” (Ps. 100:3). “Cease striving and know that I am God; I will be exalted among the nations, I will be exalted in the earth.” (Ps. 46:10).
When kids are taught about God, they typically don’t need help understanding that God exists, even though He is unseen. Thus, our desire to worship Him is not a learned behavior. Mankind’s desire to worship and address spiritual matters is part of our divine programming. It also forms the basis of God’s First, Second, and Third Commandments. (Ex. 20:1-8; Dt. 5:1-12).
(5) Mankind’s Unique Ability to Concern Itself with Morality and Justice.
As another consequence of being made in God’s image, humans have the unique ability to concern themselves with questions of morality and justice.
Humans' unique moral system. No hominid ever created a system of laws to decide disputes between right and wrong. Likewise, nothing like that exists within the animal kingdom. This is one of the many things that makes us unique.
Moral universalism. If evolution were true, mankind should have evolved with different systems of morality regarding how humans treat each other. It is true that there are substantial differences across cultures on some points. Modern cultures, for example, disagree about things like the morality of killing whales for food. Sadly, modern cultures also disagree about how men treat women. But there are basic universal morals that are shared. For example, C. S. Lewis observed that although many past (and a few current) cultures permitted polygamy, up until very recently, every human culture has recognized the sacredness of marriage. Murder is also something that is universally considered to be wrong. The same is true with theft. Lying is another example.
God’s law written in mankind’s hearts. The Bible reveals that mankind has universal morals in terms of how it treats others because God’s morality is written on people’s hearts: “For when Gentiles who do not have the Law do instinctively the things of the Law, these, not having the Law, are a law to themselves, in that they show the work of the Law written in their hearts, their conscience bearing witness and their thoughts alternately accusing or else defending them,” (Ro. 2:14-15). These universal truths regarding how people treat each other form the basis for the Fifth through Tenth Commandments. (Ex. 20:12-17; Dt. 5:16-21).
God’s desire that we be holy. To separate mankind from the savagery seen in the animal kingdom, God desires that mankind show His image within it by being holy at all times: ‘“For I am the LORD your God. Consecrate yourselves therefore, and be holy, for I am holy . . .”’ (Lev. 11:44(a); Ep. 1:4; 1 Pet. 1:16).
(6) Mankind’s Unique Ability for Governance and Stewardship.
As another consequence of being made in God’s image, humans have the unique ability to concern themselves with matters of governance. This includes both honest and fair government and being good stewards of the environment and the animal kingdom.
Mankind’s universal desire for governance. Throughout all indigenous cultures across the world, mankind has always organized itself under systems of governance. There is no example of any people in history who lived in harmony near each other without a system of government. Government is the means by which disputes are settled, rules are enforced and the means by which people are protected. By contrast, the hominids only had family clans, just like the animal kingdom. They did not govern between family clans. There was also no central authority for setting the rules, resolving disputes, and protecting their populations.
The Bible’s revelation that government comes from God. The Bible reveals God is also the source of mankind’s desire to be governed. “It is He who changes the times and the epochs; He removes kings and establishes kings; He gives wisdom to wise men and knowledge to men of understanding.” (Dan. 2:21). “Every person is to be in subjection to the governing authorities. For there is no authority except from God, and those which exist are established by God.” (Ro. 13:1).
Mankind’s unique desire to steward the environment. Humans are also unique in their ability to care for and manage other species and the environment as a whole. There is again no evidence in the fossil record that the hominids ever did this.
God’s revelation that mankind’s duty to be stewards of the environment. The Bible reveals that mankind’s desire to govern and the concern that many have for the environment also comes from God: “Then God said, ‘Let Us make man in Our image, according to Our likeness; and let them rule over the fish of the sea and over the birds of the sky and over the cattle and over all the earth, and over every creeping thing that creeps on the earth.”’ (Gen. 1:26). Mankind is arguably failing in this God-given duty. It is estimated that humans have in recent times caused the extinction rate to rise between 1,000 and 10,000 times higher than the natural extinction rate.296 In 2002, biologist E. O. Wilson estimated one-half of all plant and animal species of life on earth will be extinct in 100 years.297
source:298
(7) Mankind’s Unique Ability to Concern Itself with Love and Compassion.
Finally, as a consequence of being made in God’s image, humans have the unique ability to show love and concerns for others. This is by far His greatest charge and gift to mankind.
The difference between human and animal love and compassion. As a general rule, mammals show a form of compassion for their young. Without this compassion, their young would die. Birds also show this concern for their young. No doubt the hominids also cared for their young. Dogs can also show compassion for humans when adopted by a human family. Yet, as a general rule, neither animals nor the hominids showed a systematic love and compassion for others outside of their clans or families. Mankind is unique in its ability to show systematic love and compassion to strangers. Many fail to use this gift. But it is a gift that God meant to be used.
God created mankind for good works. In addition to fellowship with Him, God created mankind to do “good works” through acts of love and compassion toward strangers: “For we are His workmanship, created in Christ Jesus for good works, which God prepared beforehand so that we would walk in them.” (Eph. 2:10).
God created mankind to love one another. God’s greatest commandment is to love others: “This I command you, that you love one another.” (Jo. 15:17). “Now we who are strong ought to bear the weaknesses of those without strength and not just please ourselves.” (Ro. 15:1). This love requires a believer to act “with all humility and gentleness, with patience, showing tolerance for one another in love,” (Eph. 4:2). “We urge you, brethren, admonish the unruly, encourage the fainthearted, help the weak, be patient with everyone.” (1 Thess. 5:14). Your God-given ability to love others is also His greatest gift to you: “But now faith, hope, love, abide these three; but the greatest of these is love.” (1 Cor. 13:13).
Conclusion: Give thanks for your creation. The psalmist David knew that he owed his existence to God. He also knew that God created him while he was in his mother’s womb. Knowing this truth, he sang God’s praise: “I will give thanks to You, for I am fearfully and wonderfully made; wonderful are Your works, and my soul knows it very well.” (Ps. 139:14). Isaiah also revealed that God made you in your mother’s womb: “Thus says the LORD who made you and formed you from the womb, who will help you, ‘Do not fear, O Jacob My servant; and you Jeshurun whom I have chosen.”’ (Is. 44:2).
The Genesis text also reveals that God referred to Himself in the plural form: “Then God said, ‘Let Us make man in Our image, according to Our likeness; . . .’” (Gen. 1:26). The New Testament reveals that Jesus was part of the triune God who played a central role in your creation: “For by Him all things were created, both in the heavens and on earth, visible, and invisible, whether thrones or dominions or rulers or authorities -- all things have been created through Him and for Him.” (Col. 1:16). “All things came into being through Him, and apart from Him nothing came into being that has come into being.” (Jo. 1:3). “Yet for us there is but one God, the Father, from whom are all things and we exist for Him; and one Lord, Jesus Christ, by whom are all things, and we exist through Him.” (1 Cor. 8:6). “fixing our eyes on Jesus, the author and perfecter of faith, who for the joy set before Him endured the cross, despising the shame, and has sat down at the right hand of the throne of God.” (Heb. 12:2).
Jesus has revealed the truths of His creation to show that He is all powerful and that there is none like Him: “To you it was shown that you might know that the LORD, He is God; there is no other besides Him.” (Dt. 4:35). “Know therefore today, and take it to your heart, that the LORD, He is God in heaven above and on the earth below; there is no other.” (Dt. 4:39). “For since the creation of the world His invisible attributes, His eternal power and divine nature, have been clearly seen, being understood through what has been made, so that they are without excuse.” (Ro. 1:20).
As the Creator of the universe and Creator of mankind, there is no problem in your life that is too big for Him to solve. Are you turning to your Creator in faith to solve your problems? Are you giving Him praise and thanks?
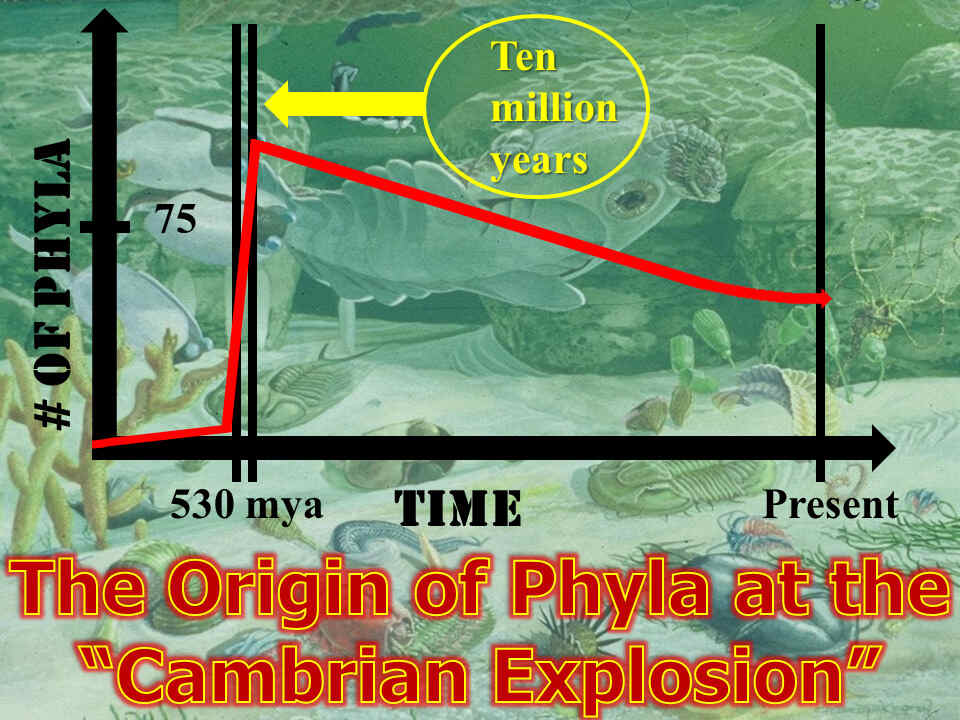
Give thanks that Jesus made you out of love for you299

Give thanks that Jesus died to offer you a path to eternal life with Him300
Source for both images: Michelangelo Buonarroti: The Creation of Adam (artbible.info)↩︎
Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; Slack, John F.; Rittner, Martin; Pirajno, Franco; O'Neil, Jonathan; Little, Crispin T. S. (1 March 2017). "Evidence for early life in Earth's oldest hydrothermal vent precipitates". Nature (journal). 543: 60–64. doi:10.1038/nature21377. Retrieved 2 March 2017; Zimmer, Carl (1 March 2017). "Scientists Say Canadian Bacteria Fossils May Be Earth's Oldest". New York Times. Retrieved 2 March 2017; Dunham, Will (1 March 2017). "Canadian bacteria-like fossils called oldest evidence of life". Reuters. Retrieved 1 March 2017.↩︎
Hugh Ross, The Creator and the Cosmos, p. 148; Niles Eldredge The Triumph of Evolution and the Failure of Creationism (W.H. Freeman and Company 2000) p.37.↩︎
Nick Lane, First breath: Earth's billion-year struggle for oxygen" New Scientist, 6 Feb. 2010, pp. 36-9.↩︎
Carl Woese, J Peter Gogarten, “When did eukaryotic cells (cells with nuclei and other internal organelles) first evolve? What do we know about how they evolved from earlier life-forms?” Scientific American, October 21, 1999.↩︎
L. Paul Knauth and Martin J. Kennedy, the Late-Precambrian Greening of the Earth,” Nature 460 (August 6, 2009), p. 728-32; cited in, Hugh Ross Navigating Genesis (rtb press 2014), p. 51.↩︎
↩︎Steven J. Gould, Wonderful Life: The Burgess Shale and the Nature of History (New York: W.W. Norton, 1989), 222-24; Simmon Coway Morris, "The Community Structure of the Middle Cambrian Phyllopod Bed (Burgess Shale)," Paleontology 29 (1986): 423-67; Hugh Ross, Navigating Genesis (rtb press 2014) p. 58.
“Evolution of amphibians". University of Waikato: Plant and animal evolution. September 30, 2012↩︎
Laurin, M.; Reisz, R. R. "A reevaluation of early amniote phylogeny". Zoological Journal of the Linnean Society (1995) 113 (2): 165–223.↩︎
Rose, Kenneth D. "The earliest primates". Evolutionary Anthropology: Issues, News, and Reviews (Hoboken, NJ: John Wiley & Sons) (1994). 3 (5): 159–173.↩︎
Fossil Reanalysis Pushes Back Origin of Homo sapiens". Scientific American (Stuttgart: Georg von Holtzbrinck Publishing Group). February 17, 2005.↩︎
Mellars, Paul, "Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model". Proc. Natl. Acad. Sci. U.S.A. (Washington, D.C.: National Academy of Sciences) (June 20, 2006). 103 (25): 9381–9386.↩︎
An example of God’s dual type of creation is His creation of Adam from the dust and Eve from Adam’s existing rib.↩︎
Valentine, James W, On the Origin of Phyla (Chicago: University Of Chicago Press 2004), p. 7. ISBN 0-226-84548-6.↩︎
↩︎Steven J. Gould, Wonderful Life: The Burgess Shale and the Nature of History (New York: W.W. Norton, 1989), 222-24; Simmon Coway Morris, "The Community Structure of the Middle Cambrian Phyllopod Bed (Burgess Shale)," Paleontology 29 (1986): 423-67.
↩︎Jean Vannier and Jun-Yuan Chen, "The Early Cambrian Colonization of Pelagic Niches Exemplified by Isoxys (Arthropoda)," Lethaia 33 (2000): 295-311; Philip W. Singor and Geeret J. Vermeji, "The Plankton and the Benthos: Origins and Early History of an Evolving Relationship," Paleobiology 20 (1994): 297-319.
"Catching the first fish" by Philippe Janvier; Nature 402:21 - 22 (1999).↩︎
↩︎Barry Cox, R.J.G. Savage, Brian Gardiner and Colin Harrison Encyclopedia of Dinosaurs & Prehistoric Creatures: A Visual Who's Who of Prehistoric Life (Simon & Schuster 2ed. 1999) p. 22.
D.-G. Shu et al., “Lower Cambrian Vertebrates from South China,” Nature 402 (November 4, 1999): 42-46; Jun-Yuan Chen, Di-Ying Huang, and Chia-Wei Li, “An Early Cambrian-like Chordate,” Nature 402 (December 2, 1999):518-22; Richard A. Kerr; “Evolution’s Big Bang Gets Even More Explosive,” Science 261 (September 3, 1993):1274-75; Richard A Bowring et al., “Calibrating Rates of Early Cambrian Evolution,” Science 261 (September 3, 1993); 12930-99; J.-Y. Chen et al., “A Possible Early Cambrian Chordate,” Nature 377 (October 26, 2002): 720-21; cited in Hugh Ross, Navigating Genesis (rtb press 2014) p. 58.↩︎
Hugh Ross, Navigating Genesis (rtb press 2014) p. 58, citing D.G. Shu et al, “An Early Cambrian Tunicate from China,” Nature 411 (May 24, 2001): 472-73; D.-G. Shu et al., “Primitive Deuterostomes from the Chengjiand Lagerstatte (Lower Cambrian, China),” Nature 414 (November 22, 2001): 419-24; D. Shu, X. Zhang, and L. Chen, “Reinterpretation of Yumnanzoon as the Earliast Known Hemichordate,” Nature 380 (April 4, 1996): 428-30.↩︎
Shen, Bing; Dong, Lin; Xiao, Shuhai; Kowalewski, Michał (2008). "The Avalon Explosion: Evolution of Ediacara Morphospace". Science 319 (5859): 81–84.↩︎
Seilacher, A. (1992). "Vendobionta and Psammocorallia: lost constructions of Precambrian evolution". Journal of the Geological Society, London 149 (4): 607–613.↩︎
Buckland, W. Geology and Mineralogy Considered with Reference to Natural Theology. Lea & Blanchard. (1841).↩︎
Darwin, The Origin of Species, (1859) p. 348.↩︎
Id., at p. 344.↩︎
Id., p. 350 (italics added).↩︎
Id., p. 351 (italics added).↩︎
I. Axelrod, “Early Cambrian Marine Fauna,” Science, Vol. 128, 4 July 1958, p. 7.↩︎
T. Neville George Professor of Geology at the University of Glasgow, “Fossils in Evolutionary Perspective,” Science Progress, Vol. 48, No. 189, January 1960, p. 5.↩︎
↩︎(Emphasis added.) J. W. Valentine, "The Evolution of Complex Animals" What Darwin Began, Laurie Godfrey (Boston: Allyn and Bacon 1985) p. 263, 267.
↩︎Stephan Bengston, Nature 345:765 (1990).
Barnes, R.S.K., P. Calow, P.J.W. Olive, D.W. Golding, J.I. Spicer. The Invertebrates: A New Synthesis 3rd Ed. (2001).↩︎
↩︎Dr. Duane Gish Evolution: The Fossils Still Say No!, p. 76, quoting Romer in his book Vertebrate Paleontology, p. 15.
↩︎Gish p. 78, quoting Romer at 52.
↩︎Gish at 78, quoting Romer at 53.
↩︎Gish at 79, quoting G. T. Todd, American Zoology 204: 757 (1980.)
↩︎Srahler, Science and Earth History, p. 316, quoted in Gish p. 79.
↩︎Strahler, Science And Earth History, p. 408, quoted in Gish at p. 80.
↩︎E. White, PROC. Linn. Soc. London 177:8 (1966), quoted in Gish p. 78.
Jeffrey S. Levinton, “The Big Bang of Animal Evolution,” Scientific American, Vol. 267, November 1992, p. 84.↩︎
↩︎(Emphasis added.) Richard Dawkins, The Blind Watchmaker (New York: W.W. Norton 1987) p. 229.
Lee, Michael S.Y., James B. Jago, Diego C. Garcia-Bellido, Gregory D. Edgecombe, James G. Gehling, John R. Paterson. 30 June 2011. Modern optics in exceptionally preserved eyes of Early Cambrian arthropods from Australia. Nature, Vol. 474, pp. 631-634.↩︎
Example image of a compound eye: Compound microscope hi-res stock photography and images - Alamy↩︎
Image credit: Opabinia (lobópodo del Cámbrico, 505MA) (Olof Helje)↩︎
Image credit: lifebeforethedinosaurs.com/2011/06/haikouichthys.html↩︎
↩︎Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p.23.
Nick Lane, First breath: Earth's billion-year struggle for oxygen" New Scientist, 6 Feb. 2010, pp.36-9.↩︎
Hugh Ross, Navigating Genesis (2014 rtb press) p. 59.↩︎
↩︎quoted in, Philip Johnson, Darwin on Trial (InterVarsity Press 2ed. 1993) p. 46.
↩︎Id. at 46-47.
↩︎Id. at 46.
↩︎(Emphasis added.) Thomas Huxley in 3 lectures on evolution (1882), 619, quoted in John Ankerberg & John Weldon, Darwin’s Leap of Faith (Harvest House 1998) p. 210.
↩︎(Emphasis added.) Stephen J. Gould, The Return of Hopeful Monsters, Natural History, (June - July 1977) pp. 22, 24, quoted in John Ankerberg & John Weldon, Darwin’s Leap of Faith (Harvest House 1998) p. 212.
Gould, S. J., "Is a new and general theory of evolution emerging?" Paleobiology, vol. 6(1), p. 119-130 (1980).↩︎
Gould, S. J., "Evolution's erratic pace," Natural History, Vol. 86, No. 5, pp. 12-16, (May 1977).↩︎
↩︎G. Simpson, The Major Features of Evolution (1943), p. 143; G. Simpson, Tempo and Mode in Evolution (1944), p. 107, G. Simpson, The Major Features of Evolution (1965) p. 360, quoted in John Ankerberg & John Weldon, Darwin’s Leap of Faith (Harvest House 1998) p. 213.
Eldredge, N., Reinventing Darwin, p. 95 (1995).↩︎
Carroll, R., Patterns and Processes of Vertebrate Evolution, pgs. 8-10 (1997).↩︎
Pagel M., "Happy accidents?," Nature, Vol 397, pg. 665 (February 25, 1999).↩︎
Mayr, E., What is Evolution, pg. 189 (2001).↩︎
Eldredge, Niles and S. J. Gould "Punctuated equilibria: an alternative to phyletic gradualism" In T.J.M. Schopf, ed., Models in Paleobiology. (San Francisco: Freeman Cooper (1972), pp. 82-115. Reprinted in N. Eldredge Time frames. Princeton: Princeton Univ. Press, 1985, pp. 193-223.↩︎
Gould, S. J., "Is a new and general theory of evolution emerging?" Paleobiology, vol 6(1), p. 119-130 (1980).↩︎
Id.↩︎
Richard Milton in Shattering the Myths of Darwinism (Park Street Press, VE 1997) p. 145-46.↩︎
Id.↩︎
Id.↩︎
↩︎(emphasis original.) Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 40.
↩︎(Emphasis original.) Id. p. 243.
↩︎(Emphasis original) Id. at 246.
↩︎Id. at 125.
Gould, S. J., "Is a new and general theory of evolution emerging?" Paleobiology, vol 6(1), p. 119-130 (1980).↩︎
↩︎(Emphasis added.) Richard Dawkins, The Blind Watchmaker (New York: W.W. Norton 1987) pp. 240- 241.
See Teaching About Evolution and the Nature of Science (National Academy Press, 1998), pg. 43, 56, 57; Science and Creationism, A View from the National Academy of Sciences, 2nd Ed (1999).↩︎
Kitzmiller v. Dover Area Sch. Dist., 400 F. Supp. 2d 707, 735-36 (M.D. Pa. 2005).↩︎
Id., at 742-3.↩︎
Stanley, S., Macroevolution, pg. 93.↩︎
↩︎(Italics original; bold added.) Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 230.
↩︎Id. at p. 169.
Teaching About Evolution and the Nature of Science (National Academy Press, 1998), pg. 43, 56, 57.↩︎
Stephen Jay Gould as quoted in Lewin, R., "Evolutionary Theory Under Fire," Science Vol 210:883-887 (November, 1980).↩︎
Eldredge, N. The Triumph of Evolution and the Failure of Creationism (2000).↩︎
↩︎Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 57.
↩︎Id. at 245.
↩︎Id.
Charlesworth, B., Lande, R., and Slatkin, M., "A Neo-Darwinian Commentary on Macroevolution," Evolution, 36(3) 1982 pg. 474-498.↩︎
↩︎P . Dobzhansky, 1951. Genetics and the Origin of Species (New York: Columbia University Press), p. 59, quoted in Of Pandas and People: The Central Question of Biological Origins, Charles Thaxton (editor) (Haughton Publishing Co. 4th ed. 1999) p. 65.
↩︎Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 57.
↩︎Id. at 125.
↩︎(Emphasis added) Id. at 247.
↩︎(Emphasis original) Of Pandas and People: The Central Question of Biological Origins, Charles Thaxton (editor) (Haughton Publishing Co. 4th ed. 1999) p. 66.
↩︎Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 73.
↩︎(emphasis added.) Id. 57.
↩︎Note: A homozygote is an individual who carries two copies of the same gene. By contrast, a heterozygote carries different copies of the same gene.
↩︎(Emphasis added.) Ian Tattersall and Jeffrey Schwartz Extinct Humans (Westview Press 2000) p. 48-49.
↩︎Dawkins p. 247.
↩︎Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 57.
↩︎Fisher, R.A., The Genetical Theory of Natural Selection (Oxford 1958) p. 84.
↩︎Dr. Lee Spetner Not by Chance: Shattering the Modern Theory of Evolution (Judaica Press 1998) p. 103.
↩︎Of Pandas and People: The Central Question of Biological Origins, Charles Thaxton (editor) (Haughton Publishing Co. 4th ed. 1999) p. 66.
↩︎(emphasis added.) Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 170. Accord Richard Dawkins The Selfish Gene (Oxford University Press 1989) p. 37 (“There is no gene which single-handedly builds a leg, long or short.”)
↩︎Charles Thaxton (editor) Of Pandas and People: The Central Question of Biological Origin (Haughton Publishing Co. 1999) p. 68 (Emphasis added), citing, E. J. Ambrose The Nature and Origin of the Biological World (New York: Wiley, Halsted 1992) p. 123.
↩︎Id.
↩︎Id.
↩︎Id.
↩︎Id.
↩︎Thaxton at 69, quoting Ambrose. at 143.
Hugh Ross, Navigating Genesis (2014 rtb press) p. 62-64.↩︎
“Exaptation” and the related term co-option describe an alleged shift in the function of a trait during a period of alleged evolution. Under this theory, a trait can allegedly evolve because it serves one particular function. Yet, under this theory, it may subsequently serve another purpose.↩︎
Ohno, S., "The notion of the Cambrian pananimalia genome," Proc. Nat. Acad. Sci. USA, Vol 93 pg. 8,475-8,478 (August 1996) (italics added).↩︎
quoted by Ankenberg & Weldon, Darwin’s Leap of Faith, (Harvest House 1998) p. 84.↩︎
↩︎Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) at p. 230.
Cowen, R., History of Life. (Blackwell Science 2002), ISBN 1-4051-1756-7.↩︎
↩︎Niles Eldredge, The Triumph of Evolution and the Failure of Creationism (W.H. Freeman and Company 2000) p. 38.
Stephen Jay Gould, “An Asteroid to Die For,” Discover, October 1989, p. 65 (italics added).↩︎
↩︎Ankerbert & Weldon p. 212-13.
↩︎Id. at 213.
Eldredge, N. and Tattersall, I., The Myths of Human Evolution, pg. 59 (1982).↩︎
David S. Woodruff, Science, pg. 717 (May 16, 1980).↩︎
↩︎Richard Dawkins The Blind Watchmaker (W.W. Norton & Company 1996) p. 230.
↩︎R.L. Carroll, Vertebrates Paleontology and Evolution (New York: W.H. Freeman and Co. 1988) p. 138, quoted in Gish p. 83.
“Evolution of amphibians". University of Waikato: Plant and animal evolution. September 30, 2012↩︎
Carroll, Robert L.; Hallam, Anthony “Patterns of Evolution, as Illustrated by the Fossil Record”. (Elsevier. Ed. 1977). pp. 405–420.↩︎
Image credit: Crossopterygians | PPT↩︎
Of Pandas and People: The Central Question of Biological Origins (Charles Thaxton (editor) (Haughton Publishing Co. 2nd ed. 1999) p. 104.↩︎
↩︎Gish at 92.
↩︎Id. at 93.
(Emphasis original) Richard Dawkins The Blind Watchmaker (W.W. Norton & Co 1996) p. 271.↩︎
quoted in Kitzmiller v. Dover Area Sch. Dist., 400 F. Supp. 2d 707, 740 (M.D. Pa. 2005) (italics added).↩︎
↩︎Dr. Michael Denton Evolution: A Theory in Crisis (Adler & Adler, Publishers, Inc. 1996) p. 290 (italics added).
↩︎Denton, p. 290-291.
Laurin, M.; Reisz, R. R. "A reevaluation of early amniote phylogeny". Zoological Journal of the Linnean Society (1995) 113 (2): 165–223.↩︎
R. L. Paton, T. R. Smithson and J. A. Clack, "An amniote-like skeleton from the Early Carboniferous of Scotland", (abstract), Nature 398, 508-513 (8 April 1999); Monastersky, R., “Out of the Swamps, How early vertebrates established a foothold—with all 10 toes—on land.” Science News, (1999) Vol. 155, No. 21, p. 328; Chapter 6: "Walking with early tetrapods: evolution of the postcranial skeleton and the phylogenetic affinities of the Temnospondyli (Vertebrata: Tetrapoda)." In: Kat Pawley (2006). "The postcranial skeleton of temnospondyls (Tetrapoda: temnospondyli)”↩︎
Falcon-Lang, H.J.; Benton, M.J.; Stimson, M. "Ecology of early reptiles inferred from Lower Pennsylvanian trackways", Journal of the Geological Society (2007) 164 (6): 1113–1118.↩︎
Palmer, D., The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions. (ed. 1999). p. 62.↩︎
Robert L. Carroll, Vertebrate Paleontology and Evolution, (W. H. Freeman and Co., New York, 1988), p. 198↩︎
Robert L. Carroll, Patterns and Processes of Vertebrate Evolution, (Cambridge University Press, 1997), pp. 296-97.↩︎
↩︎Stephen Jay Gould, "Eight (or Fewer) Little Piggies," Natural History, vol. 100, no. 1, January 1991, p. 25.
Image credit: Do Frogs Have Amniotic Eggs?↩︎
↩︎Michael Denton Evolution: A Theory in Crisis (Adler & Adler, Publishers, Inc. 1996) p. 218-19.
↩︎Charles Thaxton (editor) Of Pandas and People: The Central Question of Biological Origins (Haughton Publishing Co. 1998) p. 68, citing E. J. Ambrose, The Nature and Origin of the Biological World (New York: Wiley, Halsted 1982) p. 26.
http://www.dailymail.co.uk/sciencetech/article-1035328/Scientists-discover-worlds-bird-lived-235million-years-ago.html#ixzz3kuedz8P7; Stein, K., Palmer, C., Gill, P.G., and Benton, M.J. "The aerodynamics of the British Late Triassic Kuehneosauridae." Palaeontology, (2008). 51(4): 967-981.↩︎
von Meyer, C.E. Hermann. "Archaeopteryx lithographica (Vogel-Feder) und Pterodactylus von Solnhofen". Neues Jahrbuch für Mineralogie, Geologie und Paläontologie (in German) (1861) 1861: 678–679.↩︎
Owen, Richard. "On the Archeopteryx of von Meyer, with a description of the fossil remains of a long-tailed species, from the lithographic stone of Solenhofen". Philosophical Transactions of the Royal Society of London (1863). 153: 33–47.↩︎
Huxley, T.H. "On the animals which are most nearly intermediate between birds and reptiles". Annals and Magazine of Natural History. (1868). 4th 2: 66–75; Huxley, Thomas H. "Further evidence of the affinity between the dinosaurian reptiles and birds". Quarterly Journal of the Geological Society of London 26 (1-2): 12–31 (1870).↩︎
Image credit: Archaeopteryx - It's Nature↩︎
Seeley, Harry G. Dragons of the Air: An Account of Extinct Flying Reptiles. London: Methuen & Co. (1901) p. 239.↩︎
Reece, J, Meyers, N, Urry, L, Cain, M, Wasserman, S, Minorsky, P, Jackson, R, Cooke, B,. Cambell Biology, 9th Edition. Pearson. p. 472.↩︎
Norell, M & Ellison M Unearthing the Dragon, The Great Feathered Dinosaur Discovery (2005) Pi Press, New York,↩︎
Jerry A. Coyne., Why Evolution Is True. (Viking, New York, 2009), pgs. 195-6.↩︎
↩︎Quoted by Robert Lee Hotz, Los Angeles Times (June 23, 2000) A16.
↩︎Storrs Olson Smithsonian ornithologist, quoted by John Noble Willford The San Diego Union-Tribune (June 23, 2000) A2.
Novacek, M.J. 1994. The Radiation of Placental Mammals, in Prothero, D.R. and Schoch, R.M. (editors) Major Features of Vertebrate Evolution. Paleontological Society Short Courses in Paleontology, No. 7, pp. 220-237.↩︎
Eldredge, N., The Monkey Business: A Scientist Looks at Creationism, pg. 65-66, (1982).↩︎
Cooper, A., and Fortey, R. "Evolutionary Explosions and the Phylogenetic Fuse" Tree vol 13, no 4 (1998).↩︎
↩︎Gish at 188.
↩︎Barry Cox, R.J.G. Savage, Brian Gardiner and Colin Harrison Encyclopedia of Dinosaurs & Prehistoric Creatures: A Visual Who's Who of Prehistoric Life (Simon & Schuster 2ed. 1999), p. 198.
↩︎Cox, et al., p. 198-99.
↩︎Cox, et al., p 199.
↩︎Gish at 169.
↩︎Gish at 169-170.
↩︎Kemp at p. 271, quoted by Gish at 170.
↩︎Gish at 170.
↩︎Gish at 171.
Dr. Michael Denton Evolution: A Theory in Crisis (Adler & Adler, Publishers, Inc. 1996) p. 105-7.↩︎
↩︎Quoted in Gish at 158.
↩︎Dr. Michael Denton Evolution: A Theory in Crisis (Adler & Adler, Publishers, Inc. 1996) p. 106.
↩︎Id. at p. 146.
↩︎Percival Davis and Dean Kenyon, Of Pandas and People (Haughton Publishing Company 2ed 1999) p. 13.
↩︎Gould, 1981 "Fact and Theory" article quoted in Philip E. Johnson, Darwin on Trial (InterVarsity Press 2ed 1993) p. 81-82.
quoted by Phillip Johnson, Defeating Darwinism, (InterVarsity Press 1997) p. 15.↩︎
(italics added) The National Center for Scientific Education, Inc., Voices for Evolution, p. 58, quoted in Ankerberg & Weldon, Darwin's Leap of Faith (Harvest House 1998) p. 231.↩︎
Image credit:How Did Humans Evolve? | HISTORY↩︎
Rose, Kenneth D. "The earliest primates". Evolutionary Anthropology: Issues, News, and Reviews (Hoboken, NJ: John Wiley & Sons) (1994). 3 (5): 159–173.↩︎
↩︎Campbell, Science 154:436 (1966), quoted by Gish at D. 214.
↩︎R.D. Martin, Natural History 91:26 (1992) quoted by Gish at p. 215.
↩︎(Emphasis added.) Robin McKie Dawn of Man: The Story of Human Evolution (Dorling Kindersley Publishing, Inc. 2000): 21.
↩︎Elwyn Simmons, Annals of the New York Academy of Science, 167:319 (1969) quoted by Gish at 213.
↩︎A.J. Kelso, Physical Anthropology, 2ed (New York: J.B. Lipincott, 1974) p. 142, quoted by Gish at 214.
↩︎R.L. Carrol, Vertebrate Paleontology and Evolution (New York: W.H. Freeman and Co., 1988) p. 464, quoted by Gish at p. 214.
↩︎Elizabeth Culotta, Science 256:1516 (1992) quoted by Gish at p. 217.
↩︎A.S. Romer, Vertebrate Paleontology, 3ed (Chicago: the University of Chicago Press, 1966) p. 221 quoted by Gish at p. 217.
↩︎Kelso, Physical Anthropology, p. 150, quoted by Gish at p. 217.
↩︎Martin, Primate Origins and Evolution, p. 66, quoted by Gish at p. 218.
↩︎A.J. Kelso, Physical Anthropology, p. 151, quoted by Gish at p. 219.
↩︎R.D. Martin, Primate Origins and Evolution, p. 68. quoted by Gish at p. 219.
↩︎Gish at p. 216.
↩︎Lyell Watson, Science Digest, 90:40 (1982), cited by Gish at 224.
↩︎R.D. Martin, Primate Origins and Evolution (New Jersey: Princeton University, 1990) p. 82.), quoted by Gish at 224-25.
↩︎J.S. Jones and Esta Rouhani, "How Small Was the Bottleneck?" Nature, February 6,1996, p. 448, quoted by Ankerberg and Weldon.
Fazale Rana with Hugh Ross, Who was Adam? (NavPress 2005) p. 15.↩︎
Jerry A. Coyne., Why Evolution Is True. (Viking, New York, 2009), pgs. 210-211.↩︎
↩︎R.E. Benveniste and D.J. Todaro, Nature 261:101 (1976) cited by Gish at p. 220.
↩︎J.H. Schwartz, Nature 308:501-05 (1984), quoted by Gish by 221.
Terrence W. Deacon, “Impressions of Ancestral Brains,” p. 116, ; quoted by Fazale Rana with Hugh Ross, Who was Adam? (NavPress 2005) p. 151.↩︎
↩︎Oxnard, Nature 258:389, 394 (1979) cited by Gish at 239-40.
↩︎C. E. Oxnard, The Order of Man (Newhaven: Yale University Press 1984); p. 332, cited by Gish at 240.
↩︎Zuckerman, Beyond the Ivory Tower (New York: Taplinger 1970) p. 77, quoted by Gish at p. 239.
↩︎Stem and Susman, American Joumal of Physical Anthropology 60:279 (1983) p. 284, quoted by Gish at 251-52.
↩︎Robin McKie Dawn of Man: The Story of Human Evolution (2000 Dorling Kindersley Publishing, Inc.): 111.
↩︎Tattersall and Schwartz, Extinct Humans (WestView Press 2000) p. 90-91.
Brian G. Richmond and David S. Strait, “Evidence That Humans Evolved from a Knuckle-Walking Ancestor,” Nature 404 (2000): 382-385; Mark Collard and Leslie C. Aiello, “From Forelimbs to Two Legs,” Nature 404 (2000):339-340.↩︎
Robert Boyd and Joan B. Silk, How Humans Evolved, 3rd ed. (New York: Norton 2003), p. 282-302.↩︎
↩︎(Emphasis added) Tattersall and Schwartz, Extinct Humans (WestView Press 2000) p. 116.
↩︎Gould, 1981 "Fact and Theory" article quoted in Philip E. Johnson, Darwin on Trial (InterVarsity Press 2ed 1993) p. 81-82.
↩︎Tattersall and Schwartz, Extinct Humans (WestView Press 2000) p. 111.
↩︎Id. at 108.
↩︎R.E. F. Leakey and Alan Walker, Science 207:1103 (1980) quoted by Gish at p. 270.
↩︎Nature 261:541 (1976) Gish at 270.
Robert Boyd and Joan B. Silk, How Humans Evolved, 3rd ed. (New York: Norton 2003), p. 294-97.↩︎
↩︎S.J. Gould, Natural History, 85:30 (1976).
↩︎Robin McKie Dawn of Man: The Story of Human Evolution (2000 Dorling Kindersley Publishing, Inc.): 61.
Robert Boyd and Joan B. Silk, How Humans Evolved, 3rd ed. (New York: Norton 2003), p. 315-338; Roger Lewin, Principles of Human Evolution: A Core Textbook (Malden, MA Blackwell Science 1998) p. 309-321.↩︎
↩︎Gould, 1981 "Fact and Theory" article quoted in Philip E. Johnson, Darwin on Trial (InterVarsity Press 2ed 1993) p. 81-82.
↩︎Tattersall and Schwartz, Extinct Humans (WestView Press 2000) p. 164.
↩︎Robin McKie Dawn of Man: The Story of Human Evolution (2000 Dorling Kindersley Publishing, Inc.): 73.
Image credit: Nariokotome Boy by TheOnyxSwami on DeviantArt↩︎
↩︎Robin McKie Dawn of Man: The Story of Human Evolution (2000 Dorling Kindersley Publishing, Inc.): 79, 82.
↩︎Id. at 83.
↩︎Robin McKie Dawn of Man: The Story of Human Evolution (2000 Dorling Kindersley Publishing, Inc.): 115.
Robert Boyd and Joan B. Silk, How Humans Evolved, 3rd ed. (New York: Norton 2003), p. 294-97.↩︎
Homo heidelbergensis : Evolutionary Tree information". Smithsonian National Museum of Natural History. Retrieved 18 March 2013; Mounier, Aurélien; Marchal, François; Condemi, Silvana "Is Homo heidelbergensis a distinct species? New insight on the Mauer mandible". Journal of Human Evolution (2009). 56 (3): 219–46.↩︎
↩︎Robin McKie Dawn of Man: The Story of Human Evolution (2000 Dorling Kindersley Publishing, Inc.): 118-19.
↩︎Id. at 128.
↩︎Tattersall and Schwartz, Extinct Humans (WestView Press 2000) p. 164.
Helmuth H "Body height, body mass and surface area of the Neanderthals". Zeitschrift Für Morphologie Und Anthropologie (1998) 82 (1): 1–12.↩︎
“Neanderthal brains focused on vision and movement leaving less room for social networking”. Science Daily. March 19, 2013.↩︎
Skinner, A., B. Blackwell, R. Long, M.R. Seronie-Vivien, A.-M. Tillier and J. Blickstein; New ESR dates for a new bone-bearing layer at Pradayrol, Lot, France; Paleoanthropology Society March 28, 2007.↩︎
Finlayson, C; Pacheco, Fg; Rodríguez-Vidal, J; Fa, Da; Gutierrez, López, Jm; Santiago, Pérez, A; Finlayson, G; Allue, E; Baena, Preysler, J; Cáceres, I; Carrión, Js; Fernández, Jalvo, Y; Gleed-Owen, Cp; Jimenez, Espejo, Fj; López, P; López, Sáez, Ja; Riquelme, Cantal, Ja; Sánchez, Marco, A; Guzman, Fg; Brown, K; Fuentes, N; Valarino, Ca; Villalpando, A; Stringer, Cb; Martinez, Ruiz, F; Sakamoto, T "Late survival of Neanderthals at the southernmost extreme of Europe". Nature (October 2006) 443 (7113): 850–3; Outside Europe, Mousterian tools were made by both Neanderthals and early modern Homo sapiens. (Donald Johanson & Blake Edgar (2006) From Lucy to Language, Simon & Schuster, p. 272.↩︎
Wilford, John Noble "Neanderthals and the Dead". New York Times (December 16, 2013).↩︎
Ovchinnikov, Iv; Götherström, A; Romanova, Gp; Kharitonov, Vm; Lidén, K; Goodwin, W "Molecular analysis of Neanderthal DNA from the northern Caucasus". Nature (March 2000) 404 (6777): 490–3; Matthias Krings et al., "A View of Neanderthal Genetic Diversity," Nature Genetics 26 (2000): 144-46; Igor V. Ovchinnikov et al., "Molecular Analysis of Neanderthal DNA from the North Caucasus," Nature 404 (2000): 490-93; Matthias Krings et al., "Neanderthal DNA Sequences and the Origin of Modern Humans," Cell 90 (1997): 19-30.↩︎
“Neanderthal Genome Sequencing Yields Surprising Results And Opens A New Door To Future Studies" (Press release). Lawrence Berkeley National Laboratory. 16 November 2006.↩︎
Hayes, Jacqui "DNA find deepens Neanderthal mystery" (15 November 2006) Cosmos.↩︎
Evans PD, Mekel-Bobrov N, Vallender EJ, Hudson RR, Lahn BT "Evidence that the adaptive allele of the brain size gene microcephalin introgressed into Homo sapiens from an archaic Homo lineage". Proceedings of the National Academy of Sciences (November 2006) 103 (48): 18178–83; Evans PD, Gilbert SL, Mekel-Bobrov N, Vallender EJ, Anderson JR, Vaez-Azizi LM, Tishkoff SA, Hudson RR, Lahn BT "Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans". Science (September 2005) 309 (5741): 1717–20.↩︎
“Genome sequence of a 45,000-year-old modern human from western Siberia". Nature (23 October 2014) 514 (7523): 445–449.↩︎
The New World Dictionary of the American Language (Simon and Schuster 2 ed, 1980) p. 1367.↩︎
Hugh Ross, Navigating Genesis (rtb press 2014) p. 77.↩︎
Id.↩︎
↩︎Robin McKie Dawn of Man: The Story of Human Evolution (2000 Dorling Kindersley Publishing, Inc.): 79, 82.
↩︎Id. at 83.
↩︎John W. Oller, Jr. & John L. Omdahl ``Origin of the Human Language Capacity: In Whose Image?” The Creation Hypothesis: Scientific Evidence for An Intelligent Designer (J. P. Moreland (editor) InterVarsity Pres (1994) p. 235.
↩︎Id. at p. 255.
↩︎Id. at p. 259, citing Claire Russell and W. M. S. Russell “Language and Animal Signals” (1971) in Language: Introductory Readings (New York: St. Martin’s, 1981) (p. 233.
↩︎Id. at p. 260.
↩︎Id. at 260.
↩︎Id. at p. 262.
↩︎Id. at p. 264.
Fazale Rana with Hugh Ross, Who was Adam? (NavPress 2005) p. 85, citing Richard G. Klein, The Human Career: Human Biological and Cultural Origins, 2nd ed. (Chicago: University of Chicago Press, 1999), p. 520-529; Olga Soffer, “Late Paleolithic,” in Encyclopedia of Human Evolution and Prehistory, ed. Eric Delson et al., (New York: Garland, 2000), p. 375-80; Alison S. Brooks, “Late Stone Age,” in Encyclopedia of Human Evolution and Prehistory, p. 380-82.↩︎
Rana, et. al., Who was Adam? p. 86.↩︎
Rana, et. al., p. 87, citing, Soffer, p. 375-80.↩︎
Gardner, Helen, and Fred S. Kleiner. Gardner's Art through the Ages: A Global History. 13th ed. Australia: Thomson/Wadsworth, 2009., p. 6.↩︎
Rana, et. al., p. 88, citing, Roger Lewin, Principles of Human Evolution: A Core Textbook (Malden, MA: Blackwell Science, 1998), 384-442.↩︎
Rana, et. al., p. 89, citing Tim Appenzeller, “Evolution or Revolution?” Science 282 (1998): 1451-1454.↩︎
Rana, et. al., p. 90.↩︎
Fossil Reanalysis Pushes Back Origin of Homo sapiens". Scientific American (Stuttgart: Georg von Holtzbrinck Publishing Group). February 17, 2005.↩︎
Rana, et. al., p. 78.↩︎
Hugh Ross, Navigating Genesis (rtb press 2014) p. 75.↩︎
Shaw, Kate "Sheer Numbers Gave Early Humans Edge Over Neanderthals". (July 29, 2011) Wired.com; Vergano, Dan "Neanderthals Lived in Small, Isolated Populations, Gene Analysis Shows". (April 22, 2014). National Geographic.↩︎
Rana, et. al., p. 86.↩︎
Rana, et. al., p. 90-91.↩︎
{kind=link}